VIGS: Harnessing RNA-Mediated Reverse Genetics for Gene Function and Drug Discovery
Virus-induced gene silencing (VIGS) is a powerful RNA-mediated reverse genetics technology that enables rapid, transient knockdown of gene expression by exploiting the plant's innate RNA interference (PTGS) machinery.
VIGS: Harnessing RNA-Mediated Reverse Genetics for Gene Function and Drug Discovery
Abstract
Virus-induced gene silencing (VIGS) is a powerful RNA-mediated reverse genetics technology that enables rapid, transient knockdown of gene expression by exploiting the plant's innate RNA interference (PTGS) machinery. This article provides a comprehensive resource for researchers and drug development professionals, detailing the foundational mechanisms of VIGS, from dsRNA processing and siRNA formation to systemic silencing. It covers advanced methodological applications across diverse species, including crops and medicinal plants, and offers practical troubleshooting strategies for optimizing silencing efficiency. By comparing VIGS to other functional genomics tools and validating its role in identifying genes for stress tolerance and valuable metabolic pathways, this review underscores VIGS's transformative potential in accelerating gene discovery and informing therapeutic development.
The Core Machinery of VIGS: From Antiviral Defense to Reverse Genetics Powerhouse
Virus-induced gene silencing (VIGS) is an RNA-mediated reverse genetics technology that has evolved into an indispensable approach for analyzing gene function in plants [1]. This technique exploits the innate antiviral defense mechanism of plants, leading to sequence-specific degradation of target mRNAs. As a form of post-transcriptional gene silencing (PTGS), VIGS provides a powerful platform for functional genomics, enabling researchers to characterize gene function without the need for stable transformation [2]. The methodology has transformed plant molecular biology by offering a rapid, efficient, and specific system for transient gene silencing, making it particularly valuable for species recalcitrant to genetic transformation [3] [4].
Molecular Mechanisms of VIGS
The biological basis of VIGS lies in the plant's natural RNA silencing machinery, which is typically deployed as a defense mechanism against viral pathogens [3]. The process represents a sophisticated RNA-mediated defense system that can be harnessed for experimental gene silencing.
Core Mechanism
The VIGS process initiates when a recombinant viral vector containing a fragment of a host target gene is introduced into the plant [1]. The molecular cascade unfolds through these critical steps:
- Vector Transcription: The T-DNA carrying the viral genome is transformed into the plant by Agrobacterium and transcribed by the host's RNA polymerase II [5].
- dsRNA Formation: Viral RNA-dependent RNA polymerase (RdRP) produces double-stranded RNA (dsRNA) from the single-stranded RNA (ssRNA) viral transcripts [1] [5].
- dicer Processing: Cellular Dicer-like (DCL) enzymes recognize and cleave these dsRNAs into small interfering RNA (siRNA) duplexes of 21–24 nucleotides [1] [3].
- RISC Assembly: These siRNAs are incorporated into the RNA-induced silencing complex (RISC), where the complex uses the siRNA as a guide to identify complementary mRNA sequences [1] [5].
- Target Degradation: The Argonaute (AGO) protein, a core component of RISC, enables sequence-specific cleavage and degradation of the target mRNA, resulting in gene silencing [1].
This process is depicted in the following workflow diagram:
Systemic Silencing and Amplification
A crucial feature of VIGS is its systemic nature. The silencing signal amplifies and spreads throughout the plant via secondary siRNA production [1]. Mobile silencing signals, likely involving the 21-nucleotide siRNA class, facilitate this systemic propagation, enabling gene silencing in tissues distant from the initial infection site [3]. This systemic movement is particularly dependent on the phloem, allowing the signal to reach various organs and produce whole-plant phenotypes [6].
VIGS Vector Systems
The effectiveness of VIGS depends significantly on the viral vector system employed. Different vectors offer distinct advantages and limitations based on their host range, symptom severity, and capacity for foreign DNA insertion.
Major Vector Classes
VIGS vectors are broadly categorized into three classes based on their viral origin: RNA viruses, DNA viruses, and satellite virus-based systems [3]. To date, approximately 50 viral vectors have been developed for VIGS applications across diverse plant species [3].
Table 1: Major VIGS Vector Systems and Their Applications
| Vector Name | Virus Type | Host Range | Key Features | Applications |
|---|---|---|---|---|
| Tobacco Rattle Virus (TRV) | RNA virus | Broad (Solanaceae, Cruciferae, Gramineae) | Mild symptoms, meristem invasion, efficient systemic movement [3] [5] | Most widely used system; functional genomics in model plants and crops [3] [7] |
| Tobacco Mosaic Virus (TMV) | RNA virus | Moderate | First VIGS vector developed [1] | Initial proof-of-concept studies [1] |
| Bean Pod Mottle Virus (BPMV) | RNA virus | Soybean | Efficient in legumes [7] | Soybean functional genomics [7] |
| Barley Stripe Mosaic Virus (BSMV) | RNA virus | Monocots (barley, wheat) | Effective in cereal crops [2] | Gene function studies in monocots [2] |
| Geminiviruses (CLCrV, ACMV) | DNA virus | Variable | Nuclear replication, potential for epigenetic studies [3] | Specialized applications in specific hosts [3] |
The TRV Vector System
The TRV-based VIGS system has emerged as one of the most versatile and widely adopted platforms, particularly for solanaceous plants [3]. Its bipartite genome organization requires two plasmid components:
- TRV1: Encodes replicase proteins for viral replication, movement protein for cell-to-cell movement, and a weak RNA silencing suppressor [3].
- TRV2: Contains the coat protein gene and a multiple cloning site (MCS) for inserting target gene fragments [3].
The development of advanced TRV vectors featuring GATEWAY cloning technology or ligation-independent cloning has significantly streamlined the construction of silencing vectors, enhancing throughput and efficiency [5].
Experimental Design and Optimization
Implementing a robust VIGS protocol requires careful consideration of multiple experimental parameters that significantly influence silencing efficiency.
Critical Experimental Parameters
The following factors are crucial for successful VIGS experiments:
Insert Fragment Design: Optimal fragment sizes range from 200 to 500 base pairs with high sequence specificity to avoid off-target effects [2] [5]. The insert should lack homopolymeric regions and possess high gene specificity [5].
Plant Developmental Stage: Younger plants typically show higher silencing efficiency, with the optimal stage being 2-4 weeks post-germination for many species [3].
Environmental Conditions: Temperature significantly impacts silencing efficiency, with 20-22°C being optimal for many systems post-inoculation [3] [6]. Light intensity and humidity also influence outcomes.
Agroinoculum Concentration: Optical density (OD₆₀₀) typically ranges from 0.5 to 2.0, with species-specific optimization required [3] [4].
Table 2: Quantitative Parameters for VIGS Optimization
| Parameter | Optimal Range | Impact on Silencing | References |
|---|---|---|---|
| Insert Size | 200-500 bp | Larger fragments may reduce efficiency; smaller fragments may lack specificity [2] [5] | [2] [5] |
| Agroinoculum OD₆₀₀ | 0.5-2.0 | Species-dependent; affects infection efficiency and symptom severity [3] [4] | [3] [4] |
| Post-inoculation Temperature | 20-22°C | Lower temperatures generally enhance silencing spread and durability [3] [6] | [3] [6] |
| Time to Phenotype | 2-4 weeks | Varies by species, target gene, and vector system [4] [5] | [4] [5] |
| Silencing Efficiency | 65-95% | Dependent on optimization of all parameters [7] | [7] |
Advanced Optimization Strategies
Recent advances have focused on enhancing VIGS efficiency through molecular optimization. A particularly promising approach involves engineering viral suppressors of RNA silencing (VSRs). For example, a truncated version of the Cucumber Mosaic Virus 2b protein (C2bN43) retains systemic silencing suppression while losing local suppression activity, resulting in significantly enhanced VIGS efficacy in pepper [6].
Applications in Functional Genomics
VIGS has been successfully applied to characterize genes involved in diverse biological processes, significantly accelerating functional genomics in numerous plant species.
Applications Across Plant Species
The technology has expanded beyond model plants to encompass a broad range of species:
- Model Plants: Nicotiana benthamiana and Arabidopsis thaliana remain primary systems for VIGS methodology development [3].
- Solanaceous Crops: Tomato, pepper, and potato have seen extensive application of VIGS for studying disease resistance, fruit development, and abiotic stress tolerance [3].
- Monocot Species: BSMV-based vectors enable VIGS applications in barley, wheat, and other cereals [2].
- Horticultural Species: Recent advances have established VIGS in ornamental species like Agapanthus praecox for studying flower coloration [4].
- Legumes: Soybean VIGS systems have been developed using both BPMV and TRV vectors [7].
Functional Characterization of Specific Processes
VIGS has enabled significant advances in understanding plant biology:
- Biotic Stress Resistance: VIGS has identified numerous resistance (R) genes and components of defense signaling pathways [3] [7].
- Abiotic Stress Tolerance: Genes involved in drought, salinity, and temperature stress responses have been characterized using VIGS [1] [3].
- Metabolic Pathways: VIGS has elucidated regulatory genes controlling specialized metabolism, including anthocyanin biosynthesis in Agapanthus [4] and capsaicinoid biosynthesis in pepper [3].
- Plant Development: Genes regulating architecture, flowering, and meristem function have been successfully characterized through VIGS approaches [3].
Advanced Applications and Future Directions
VIGS in Medicinal Plants and Drug Discovery
VIGS represents a particularly valuable tool for medicinal plant research, where complex genomes and secondary metabolite pathways present significant challenges [8]. With over 400 medicinal plant genomes sequenced as of 2025, VIGS enables functional characterization of genes involved in the biosynthesis of valuable pharmaceutical compounds [8]. This application accelerates the identification of key enzymes in pathways producing therapeutic agents such as vinblastine (Catharanthus roseus), artemisinin (Artemisia annua), and paclitaxel (Taxus brevifolia) [8].
VIGS-Induced Epigenetic Modifications
Emerging evidence indicates that VIGS can induce heritable epigenetic modifications in plants [1]. When the viral vector insert corresponds to promoter sequences rather than coding regions, VIGS can trigger RNA-directed DNA methylation (RdDM), leading to transcriptional gene silencing [1]. This epigenetic silencing can persist transgenerationally, with studies demonstrating stable inheritance of silencing phenotypes over multiple generations [1]. This application expands VIGS beyond traditional PTGS into the realm of epigenetic research and breeding.
Integration with Emerging Technologies
The future of VIGS lies in its integration with other advanced technologies:
- CRISPR Integration: Combining VIGS with CRISPR-dCas9 systems for precise epigenetic modifications [1].
- Multi-Omics Approaches: Correlating VIGS phenotypes with transcriptomic, proteomic, and metabolomic profiles for comprehensive functional analysis [3] [6].
- High-Throughput Screening: Development of VIGS-based platforms for large-scale functional screening in non-model species [3].
The Scientist's Toolkit: Essential Research Reagents
Successful implementation of VIGS requires specific reagents and materials optimized for different experimental needs.
Table 3: Essential Research Reagents for VIGS Experiments
| Reagent/Material | Function/Purpose | Examples/Specifications |
|---|---|---|
| Binary VIGS Vectors | Delivery of target gene fragments into plant cells | pTRV1/pTRV2 systems; GATEWAY-compatible versions [3] [5] |
| Agrobacterium Strains | Mediate plant transformation | GV3101, LBA4404; optimized for virulence [4] [7] |
| Selection Antibiotics | Maintain plasmid stability in bacterial and plant systems | Kanamycin, rifampicin; concentration varies by vector system [6] [7] |
| Induction Agents | Activate virulence genes in Agrobacterium | Acetosyringone; typically 100-200 μM in infiltration medium [4] [7] |
| Infiltration Buffers | Maintain Agrobacterium viability during inoculation | MS salts, MES buffer, sugars; pH ~5.5 [4] [7] |
| Marker Genes | Visual assessment of silencing efficiency | PDS (photo-bleaching), GST (anthocyanin reduction) [4] [7] |
| VSR Enhancers | Increase silencing efficiency in recalcitrant species | Modified viral suppressors (e.g., C2bN43) [6] |
Virus-induced gene silencing represents a sophisticated integration of molecular biology and plant pathology, harnessing the plant's innate antiviral defense for targeted gene function analysis. As outlined in this technical guide, VIGS has evolved from a simple tool for gene knockdown to a versatile platform enabling high-throughput functional genomics, epigenetic modification, and metabolic engineering. The continued refinement of vector systems, optimization protocols, and integration with emerging technologies ensures that VIGS will remain a cornerstone technique in plant molecular biology, with expanding applications in crop improvement, drug discovery, and basic research.
Virus-induced gene silencing (VIGS) is an RNA-mediated reverse genetics technology that has evolved into an indispensable approach for analyzing gene function in eukaryotes [1]. This process leverages the plant's innate antiviral defense mechanism, which operates through a sequence-specific RNA silencing pathway triggered by double-stranded RNA (dsRNA) [9]. The core of this defense is the RNA-induced silencing complex (RISC), a multiprotein complex that functions as a key tool in gene regulation at both transcriptional and translational levels [10]. Understanding the precise molecular journey from viral dsRNA detection to RISC-mediated degradation of complementary mRNA is fundamental for advancing VIGS technology and its applications in crop improvement and therapeutic development [11] [12]. This technical guide delineates these core mechanisms within the context of modern VIGS research.
The Core Mechanism: From Viral dsRNA to mRNA Cleavage
The antiviral RNAi pathway is a conserved defense mechanism that restricts viral invasion by cleaving viral RNA transcripts and repressing viral protein translation [13]. The process begins when viral RNAs and their replication intermediates form dsRNA structures within the infected plant cell.
The following diagram illustrates the complete sequence of events from viral infection to mRNA degradation, integrating the key components and processes involved in the canonical pathway.
Key Stages of the RNAi Pathway
1. Initiation: Viral dsRNA Recognition and Processing The process initiates when the RNase III enzyme Dicer (or its plant homologs, DICER-like proteins, DCLs) recognizes and cleaves long viral dsRNA molecules into small interfering RNA (siRNA) duplexes of 21–24 nucleotides in length, characterized by 2-nucleotide 3' overhangs on each strand [10] [13]. In plants, this step is part of the post-transcriptional gene silencing (PTGS) branch of RNAi [1].
2. Effector Complex Assembly: RISC Loading and Activation The siRNA duplex is loaded into the RNA-induced silencing complex (RISC). During this process, the duplex is unwound, and the passenger strand is degraded. The selected guide strand, which has the less thermodynamically stable 5' end, is incorporated into the core RISC component, an Argonaute (AGO) protein [10]. In Drosophila and other organisms, AGO2 is the primary Argonaute protein responsible for the "slicer" activity of RISC [14] [10].
3. Target Recognition and Cleavage The guide RNA within the active RISC directs the complex to complementary messenger RNA (mRNA) sequences through Watson-Crick base pairing [10]. Upon finding a perfectly complementary target, the AGO2 protein catalyzes the endonucleolytic cleavage of the mRNA, often referred to as "slicing" [14] [10] [9]. This cleavage occurs between nucleotides 10 and 11 relative to the 5' end of the guide siRNA [14].
4. mRNA Fragment Degradation Following the initial endonucleolytic cleavage by RISC, the mRNA fragments are rapidly degraded by the cell's general RNA decay machinery. The resulting 5' fragment is degraded from its 3' end by the exosome, a multimeric complex of 3'-to-5' exonucleases, whose activity is regulated by the Ski complex [14]. Conversely, the 3' fragment is degraded from its 5' end by the major cytoplasmic 5'-to-3' exonuclease, XRN1 [14]. This degradation occurs without prior decapping or deadenylation of the mRNA [14].
Quantitative Data in RNAi Efficiency
Impact of siRNA Structural Features on Gene Silencing Efficacy
Table 1: Experimentally Determined Impact of siRNA Design Parameters on RNAi Efficacy in Drosophila S2 Cells [15]
| Design Parameter | Tested Conditions | Impact on Knockdown Efficiency | Optimal Design |
|---|---|---|---|
| siRNA Length | 17 nt, 19 nt, 21 nt | Efficacy drastically decreased at 17 nt; restored at 19 nt | 19 nucleotides |
| Terminal Structure | Blunt ends vs. 2-nt 3' overhangs | Overhangs demonstrated greater efficacy | 2-nt 3' overhangs |
| GC Content | Varied across sequences | Influences efficiency; extreme GC levels are suboptimal | 30-50% (mammalian algorithms) |
| Seed Region (5' Stability) | ≥4 A/U bases vs. G/C rich | Lower stability (A/U rich) enhances RISC loading and efficacy | ≥4 A/U bases in the 5' terminal region |
Comparative Efficacy of dsRNA vs. siRNA in Pest Control
Table 2: Differential RNAi Efficacy of dsRNA and siRNA in Spodoptera litura Larvae [16]
| Parameter | Double-Stranded RNA (dsRNA) | Small Interfering RNA (siRNA) |
|---|---|---|
| Gene Silencing | Did not induce significant silencing | Exhibited clear insecticidal effects |
| Impact on Larval Growth | No significant impact | Disrupted osmoregulation and impaired larval fitness |
| Processing in Midgut | Inefficient conversion to functional siRNA | Direct activity, bypasses processing requirement |
| Key Limiting Factor | Low expression of Dicer-2; rapid dsRNA degradation in gut | Not limited by Dicer-2 expression |
| Environmental Stability | Greater stability in soil conditions | Lower environmental stability |
Experimental Protocols for Key RNAi Experiments
Protocol: Assessing RNAi Efficacy Using Synthetic siRNAs in Cell Culture
This protocol is adapted from studies conducted in Drosophila S2 cells to systematically evaluate how siRNA structural features influence knockdown efficiency [15].
1. Cell Culture and Transfection:
- Cell Line: Use Drosophila S2 cells.
- Culture Conditions: Maintain cells in Schneider's Drosophila medium, supplemented with 10% heat-inactivated fetal bovine serum and 1% penicillin-streptomycin at 25°C in a non-humidified incubator without CO₂ [15].
- Transfection: For optimal growth, passage cultures every 3-4 days to maintain a cell density between 2×10⁶ and 8×10⁶ cells/mL.
2. siRNA Design and Synthesis:
- Target Selection: Select target sites on the gene of interest (e.g., the Death-related inhibitor of apoptosis 1 (Diap1) gene, which allows for apoptosis-based phenotyping).
- Sequence Design: Design multiple siRNA sequences (e.g., 9 different targets), including some that follow standard design criteria (GC content of 30-50%, ≥4 A/U bases in the seed region) and others that intentionally violate these rules for comparison [15].
- Structural Variants: Synthesize siRNA variants with different lengths (e.g., 17 nt, 19 nt, 21 nt) and terminal structures (blunt ends vs. 2-nt 3' overhangs).
3. Efficiency Measurement:
- Phenotypic Observation: For genes like Diap1, observe apoptosis under microscopy as a clear knockdown phenotype.
- Cell Survival Quantification: Quantify cell survival rates to provide a measurable endpoint for knockdown efficiency.
- Molecular Validation: Use techniques like qRT-PCR to quantify mRNA levels post-transfection, normalizing to housekeeping genes (e.g., Actin or 18S rRNA) using the ΔΔCT method [16].
Protocol: In Vivo RNAi Efficacy in Insect Larvae
This protocol outlines the procedure for feeding dsRNA and siRNA to lepidopteran larvae to assess gene silencing and insecticidal effects, as performed in Spodoptera litura [16].
1. Insect Rearing and Preparation:
- Insect Source: Maintain S. litura larvae under controlled conditions (e.g., 26 ± 1°C with a 12-h light/12-h dark cycle).
- Diet Preparation: Prepare an artificial diet containing kidney bean powder, yeast extract, wheat germ powder, agar, and essential nutrients like L-ascorbic acid and streptomycin sulfate [16].
- Larval Selection: Use second-instar larvae. Starve them for 12–24 hours before the experiment to ensure uniform intake of the treated diet.
2. dsRNA/siRNA Administration:
- Dose Preparation: Add a measured amount of dsRNA or siRNA (e.g., 3 µg) to approximately 100 mg of artificial diet for every 10 larvae [16].
- Feeding Regimen: Replace the diet daily with freshly prepared feed containing the RNA molecules for a set period (e.g., 4 days). After this treatment period, provide larvae with an untreated artificial diet ad libitum.
- Control Groups: Include control groups fed with a diet lacking dsRNA/siRNA or containing non-targeting RNA.
3. Mortality and Efficacy Assessment:
- Data Recording: Record larval mortality daily for an extended period (e.g., up to 14 days) [16].
- Molecular Analysis: To confirm gene silencing, extract total RNA from larval midguts using TRIzol reagent. Synthesize cDNA and perform qRT-PCR to measure the expression levels of the target genes (e.g., mesh or iap), normalized to reference genes [16].
- Stability Studies: To investigate dsRNA stability, extract total small RNAs using a specialized kit (e.g., mirVana miRNA isolation kit) and analyze processing and degradation via Northern blotting [16].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Reagent Solutions for Viral dsRNA and RISC Mechanism Research
| Reagent / Solution | Specific Example (from search results) | Function / Application |
|---|---|---|
| Dicer-2 Source | Recombinant Dicer-2 (Drosophila) | Processes long dsRNA into siRNA duplexes in vitro and in vivo [10] [15]. |
| AGO2 Protein | Anti-AGO2 antibodies, recombinant AGO2 | Core catalytic component of RISC; mediates "slicer" activity for mRNA cleavage [14] [10]. |
| siRNA Synthesis Kit | MEGAscript T7 Kit (Invitrogen) | In vitro synthesis of dsRNA targeting specific genes (e.g., mesh, iap) [16]. |
| Specialized siRNA | Commercially predicted and synthesized siRNA (e.g., by MDBio, Inc.) | Designed for optimal gene silencing; used to bypass Dicer processing and test efficacy directly [16] [15]. |
| RNA Isolation Kit | mirVana miRNA isolation kit (Ambion) | Isolation of total small RNAs for Northern blot analysis of siRNA stability and processing [16]. |
| cDNA Synthesis Kit | PrimeScript RT Reagent Kit (TaKaRa) | First-strand cDNA synthesis from total RNA for subsequent qRT-PCR analysis [16]. |
| qRT-PCR Master Mix | SensiFAST SYBR Hi-ROX Kit (Bioline) | Quantitative real-time PCR to measure gene expression levels and confirm silencing efficacy [16]. |
| Cell Culture Media | Schneider's Drosophila Medium (Gibco) | Maintenance and transfection of Drosophila S2 cells for in vitro RNAi assays [15]. |
The molecular pathway from viral dsRNA to RISC-mediated mRNA degradation represents a sophisticated and highly regulated cellular defense mechanism. The efficiency of this pathway is not only governed by the core components like Dicer and AGO2 but is also critically dependent on the structural features of the siRNAs, including their length, terminal overhangs, and sequence composition [15]. Furthermore, significant interspecies variations exist, as evidenced by the inefficient processing of dsRNA in lepidopterans like Spodoptera litura due to low Dicer-2 expression, which can be bypassed by direct siRNA application [16]. A comprehensive understanding of these mechanisms, from initial dsRNA sensing to the final exonucleolytic decay of cleaved fragments by XRN1 and the exosome [14], provides a robust foundation for refining VIGS technology. This knowledge is pivotal for developing more effective and specific RNAi-based strategies for crop protection and therapeutic interventions.
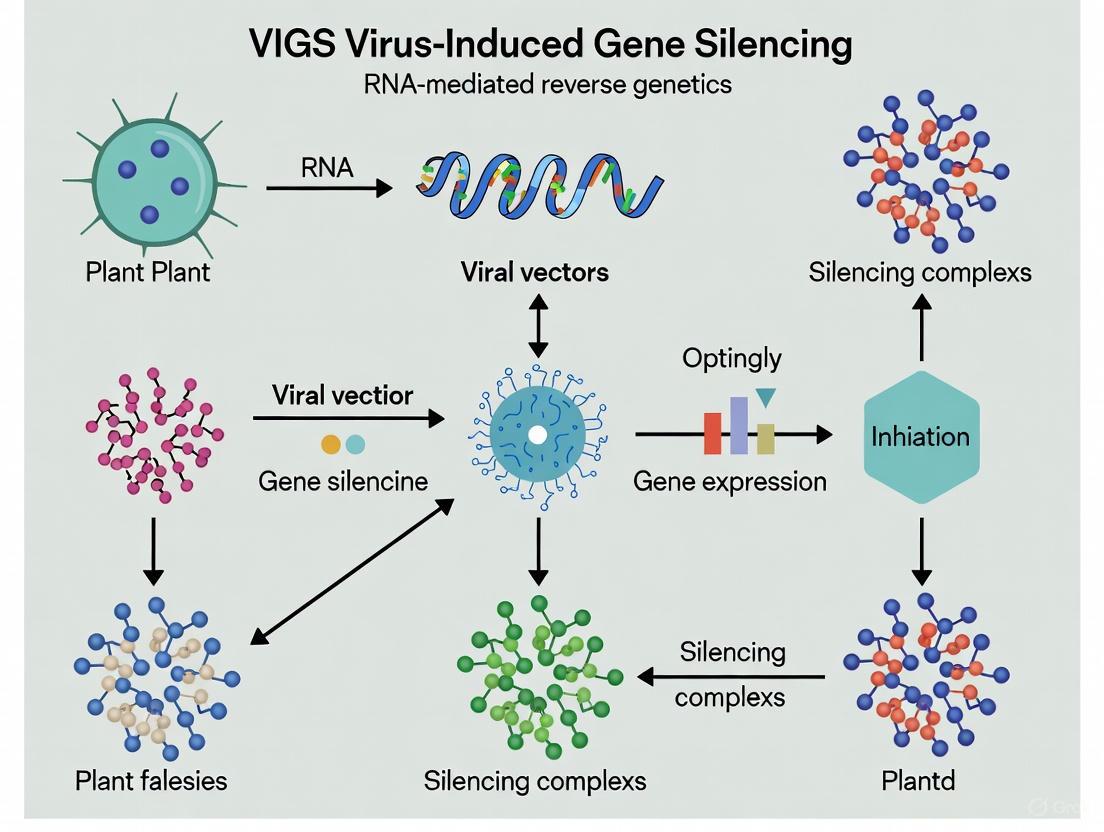
Virus-Induced Gene Silencing (VIGS) has emerged as an indispensable RNA-mediated reverse genetics technology that leverages the plant's innate antiviral defense mechanism to suppress target gene expression [1]. This powerful functional genomics tool enables researchers to characterize gene functions by introducing recombinant viral vectors carrying host gene fragments, leading to sequence-specific degradation of complementary messenger RNA (mRNA) [3] [1]. The efficacy of VIGS technology fundamentally depends on the coordinated activities of core cellular components: Dicer-like (DCL) enzymes, RNA-dependent RNA polymerase (RDRP), Argonaute (AGO) proteins, and the RNA-induced silencing complex (RISC) [3] [1] [17]. These molecular players collectively execute post-transcriptional gene silencing (PTGS), forming the backbone of not only VIGS but also broader RNA interference (RNAi) pathways across eukaryotic organisms [1] [18]. This technical guide provides a comprehensive analysis of each component's structure, function, and mechanistic role within VIGS, with particular emphasis on their applications in plant functional genomics and agricultural biotechnology.
Molecular Mechanisms of VIGS
The Core Pathway
The VIGS mechanism initiates when recombinant viral vectors introduce foreign nucleic acids into plant cells. The cellular machinery recognizes these molecules and processes them through a sophisticated multi-step pathway [3] [1]. Double-stranded RNA (dsRNA) formations, which may originate from viral replication intermediates or be synthesized by host RDRP from aberrant single-stranded RNAs, serve as the primary trigger [1] [17]. DCL enzymes then recognize and cleave these dsRNA substrates into small interfering RNA (siRNA) duplexes of defined lengths, typically 21-24 nucleotides [1] [19]. These siRNAs are loaded onto AGO proteins, the catalytic core of RISC, which subsequently unwinds the siRNA duplex and uses the guide strand to identify complementary target mRNAs [1] [18]. The activated RISC complex ultimately executes sequence-specific silencing through mRNA cleavage or translational repression [18]. In parallel, RDRP can amplify the silencing signal by synthesizing secondary dsRNA using the target mRNA as a template, thereby reinforcing and systemically spreading the gene silencing effect throughout the organism [1] [20].
Table 1: Key Steps in the VIGS Mechanism
| Step | Process | Key Players | Outcome |
|---|---|---|---|
| 1 | Trigger Recognition & dsRNA Formation | Viral Vectors, RDRP | Generation of dsRNA substrates |
| 2 | dicing | DCL Enzymes | Production of 21-24 nt siRNAs |
| 3 | RISC Loading | AGO Proteins | Formation of activated silencing complex |
| 4 | Target Recognition | RISC Complex | Identification of complementary mRNA |
| 5 | Silencing Execution | RISC Complex | mRNA cleavage or translational repression |
| 6 | Signal Amplification | RDRP | Secondary siRNA production & systemic spreading |
Visualizing the VIGS Pathway
The following diagram illustrates the coordinated sequence of molecular events during Virus-Induced Gene Silencing:
Core Cellular Components
Dicer-like (DCL) Enzymes
Dicer-like enzymes belong to the RNase III family and function as the initiating enzymes in the RNAi pathway by processing double-stranded RNA substrates into small regulatory RNAs [19] [21]. These multi-domain proteins are characterized by several conserved structural elements: DExD/H helicase domain, DUF283, PAZ domain, tandem RNase III domains, and a double-stranded RNA-binding domain (dsRBD) [19]. The PAZ domain specifically recognizes the 3' overhang of dsRNA substrates, while the RNase III domains form the catalytic core responsible for cleavage [21]. The helicase domain facilitates movement along long dsRNA molecules and contributes to substrate specificity [19].
In plants, DCL enzymes have undergone functional specialization through gene duplication and diversification. Arabidopsis thaliana possesses four DCL proteins (DCL1-DCL4) with distinct but partially overlapping functions [17]. DCL1 primarily processes hairpin-shaped precursor microRNAs (pre-miRNAs) into 21-nucleotide miRNAs that regulate endogenous gene expression [17]. DCL2 generates 22-nucleotide siRNAs from viral RNAs and endogenous transcripts, which can repress targets at the translational level and trigger amplified silencing signals [17]. DCL3 produces 24-nucleotide heterochromatic siRNAs that direct transcriptional gene silencing through RNA-directed DNA methylation (RdDM) [1]. DCL4 generates 21-nucleotide trans-acting siRNAs and the dominant class of viral siRNAs for conventional antiviral defense [17].
Table 2: DCL Family Specialization in Plants
| Enzyme | Primary Substrate | Product Size | Main Function | VIGS Relevance |
|---|---|---|---|---|
| DCL1 | pre-miRNA | 21 nt | miRNA biogenesis, development | Indirect |
| DCL2 | viral dsRNA, inverted repeats | 22 nt | Antiviral defense, translational repression | High - signal amplification |
| DCL3 | endogenous transcripts, viral DNA | 24 nt | Transcriptional silencing via RdDM | Moderate - epigenetic modifications |
| DCL4 | viral dsRNA, transgenes | 21 nt | Primary antiviral defense, tasiRNA biogenesis | High - primary antiviral response |
The critical role of specific DCL enzymes in VIGS was demonstrated in RSV CP transgenic Arabidopsis plants, where DCL2 proved essential for transgene-derived siRNA production and virus resistance, while DCL4 mutation did not compromise immunity [17]. This functional specialization highlights the sophisticated partitioning of RNA silencing pathways in plants and underscores the importance of DCL2 in particular for effective VIGS applications.
RNA-Dependent RNA Polymerase (RDRP)
RNA-dependent RNA polymerase (RdRP) represents a fundamental enzyme class that catalyzes RNA replication from an RNA template, a function contrary to the typical DNA-dependent RNA polymerases utilized for cellular transcription [20]. These enzymes are encoded in the genomes of most RNA viruses and are also found in eukaryotes, where they participate in RNAi amplification [20]. The catalytic mechanism involves a four-step process: nucleoside triphosphate (NTP) binding, active site closure, phosphodiester bond formation, and translocation [20]. Structurally, RdRps adopt a characteristic "right hand" fold composed of fingers, palm, and thumb subdomains, with the palm subdomain housing the conserved catalytic motifs (A, B, and C) responsible for coordinating metal ions and catalyzing phosphoryl transfer [22].
In VIGS, cellular RDRPs (cRdRps) serve as signal amplifiers by synthesizing secondary dsRNA using aberrant RNA molecules or cleaved target transcripts as templates [1] [20]. This activity generates additional dsRNA substrates for DCL processing, resulting in the production of secondary siRNAs that propagate and reinforce the silencing signal beyond the initial trigger [1]. This amplification mechanism enables robust and systemic silencing throughout the organism, significantly enhancing the potency and persistence of VIGS. Eukaryotic cRdRps structurally resemble simplified multi-subunit DNA-dependent RNA polymerases, particularly in their catalytic β/β' subunits, and utilize two sets of double-psi β-barrels in the active site [20].
Argonaute (AGO) Proteins
Argonaute proteins constitute the catalytic core of RNA-induced silencing complexes and represent one of the most evolutionarily conserved components of the RNAi machinery [18]. These versatile proteins directly bind small RNAs and position them for sequence-specific recognition of complementary target transcripts [18]. The eukaryotic Argonaute family divides into three principal phylogenetic clades: AGO proteins (involved in miRNA and siRNA pathways), PIWI proteins (germline-expressed, associated with piRNAs and transposon silencing), and WAGO proteins (nematode-specific Argonautes) [18].
Structurally, Argonaute proteins adopt a bilobed architecture organized around a central cleft that accommodates the guide RNA and target transcript [18]. The N-terminal lobe contains the PAZ domain, which binds the 3'-end of the small guide RNA, while the C-terminal lobe houses the MID domain (binds the 5'-phosphate of the guide) and the PIWI domain (which adopts an RNase H-like fold capable of target cleavage) [18]. The 5'-phosphate of the guide strand is anchored in a binding pocket between the MID and PIWI domains, while bases 2-8 of the guide (the "seed region") are exposed for initial target probing [18].
Beyond their canonical cytoplasmic roles in post-transcriptional regulation, AGO proteins execute multiple nuclear functions including transcriptional regulation, chromatin organization, and splicing modulation [23]. These nuclear activities expand the potential applications of VIGS beyond mRNA degradation to include epigenetic modifications. Phosphorylation at conserved residues, such as tyrosine 529 in human AGO2, represents an important regulatory mechanism that influences small RNA binding affinity and consequently modulates silencing efficacy [24].
Table 3: Argonaute Protein Classification and Functions
| Clade | Expression Pattern | Guide RNA Type | Primary Functions | Silencing Mechanism |
|---|---|---|---|---|
| AGO | Ubiquitous | miRNA, siRNA | Post-transcriptional silencing, translational repression | mRNA cleavage, translation inhibition |
| PIWI | Germline cells | piRNA | Transposon silencing, genome defense | Transcriptional silencing, mRNA degradation |
| WAGO | Nematodes | siRNA | Genome surveillance, viral defense | mRNA degradation |
RNA-Induced Silencing Complex (RISC)
The RNA-induced silencing complex represents the effector machinery of the RNAi pathway, programmed to locate and silence complementary nucleic acid targets [18]. RISC is not a single molecular entity but rather a family of heterogeneous ribonucleoprotein complexes that share a common core of Argonaute proteins bound to small regulatory RNAs [18]. These complexes range considerably in size and composition, from minimal complexes containing only an Argonaute protein and guide RNA (sufficient for target recognition and cleavage) to large "holo-RISC" particles approaching 3 MDa that incorporate numerous auxiliary factors [18].
The mechanism of RISC-mediated silencing begins with RISC loading, during which the small RNA duplex is transferred to the Argonaute protein with assistance from the Hsc70/Hsp90 chaperone machinery [19]. The duplex is then unwound, and the passenger strand is discarded, leaving the guide strand positioned for target recognition [18]. Activated RISC identifies complementary RNA transcripts through Watson-Crick base pairing with the guide RNA, with particular emphasis on perfect complementarity in the seed region (bases 2-8) [18]. Upon target engagement, RISC can silence gene expression through multiple mechanisms: endonucleolytic cleavage (catalyzed by slicer-competent AGO proteins), translational repression, or transcriptional silencing via recruitment of chromatin-modifying factors to genomic target sites [18]. The remarkable efficiency of RISC—capable of locating and cleaving targets nearly ten times faster than free RNA annealing—stems from the exposed positioning of the seed region, which functions as an initial probe while scanning the cellular transcriptome [18].
Experimental Applications in VIGS Research
Research Reagent Solutions
The following table summarizes key reagents and materials essential for implementing VIGS technology in plant systems:
Table 4: Essential Research Reagents for VIGS Implementation
| Reagent/Material | Function/Application | Examples/Specifications |
|---|---|---|
| Viral Vectors | Delivery of target gene fragments | TRV (Tobacco Rattle Virus), BBWV2, CMV, CLCrV |
| Agrobacterium Strains | Delivery of viral vectors via agroinfiltration | ABI, GV3101 |
| Plant Genotypes | Optimization of silencing efficiency | Nicotiana benthamiana, Capsicum annuum L. |
| Selection Agents | Identification of transformed plants | Glufosinate ammonium (10 mg/L) |
| Genotyping Primers | Verification of mutant plant lines | dcl2-1 LP/RP, LB1.3 for T-DNA insertion |
| Antibodies | Protein detection and silencing verification | Anti-RSV SP, Anti-myc, Anti-Rubisco-L |
VIGS Workflow for Functional Genomics
The experimental implementation of VIGS follows a systematic workflow encompassing vector design, plant inoculation, and phenotypic analysis. The following diagram outlines the key stages from vector construction to functional analysis:
Detailed Methodologies for Key Experiments
VIGS Vector Construction and Agroinfiltration
The foundational step in VIGS implementation involves cloning a 300-500 bp fragment of the target gene into a suitable viral vector, such as the bipartite Tobacco Rattle Virus (TRV) system [3]. For TRV, the fragment is inserted into the TRV2 plasmid containing the coat protein gene and multiple cloning site, while the TRV1 plasmid encodes replicase and movement proteins [3]. The recombinant vectors are then introduced into Agrobacterium tumefaciens strains (e.g., ABI) via electroporation or freeze-thaw transformation [17]. For plant inoculation, agrobacterial cultures are grown to mid-log phase (OD600 ≈ 0.4-1.0), pelleted, and resuspended in induction media (10 mM MES, 10 mM MgCl2, 150 μM acetosyringone) to final OD600 of 0.5-2.0 [3]. The bacterial suspension is infiltrated into plant tissues using needless syringes or vacuum infiltration, typically targeting expanded leaves of young plants (2-4 week stage) [17]. Critical environmental factors including temperature (18-25°C), humidity, and photoperiod must be optimized for each plant species to maximize silencing efficiency [3].
Molecular Validation of Silencing Efficiency
Comprehensive validation of successful gene silencing requires multi-level molecular analyses. Quantitative reverse-transcription PCR (qRT-PCR) provides direct measurement of target transcript reduction using gene-specific primers and reference genes (e.g., Actin or Ubiquitin) for normalization [17]. Total RNA is extracted from silenced tissues using TRIzol or similar reagents, treated with DNase I, and reverse-transcribed using oligo(dT) or random primers [17]. For protein-level validation, western blot analysis confirms reduction of the corresponding protein when specific antibodies are available [17]. Protein extracts are separated by SDS-PAGE, transferred to PVDF membranes, and probed with primary antibodies followed by HRP-conjugated secondary antibodies with chemiluminescent detection [17]. Small RNA northern blotting or high-throughput sequencing validates the accumulation of target-specific siRNAs (21-24 nt) using labeled probes complementary to the inserted fragment [17].
Genetic Approaches for Mechanism Elucidation
Determining the specific contributions of individual DCL and AGO proteins to VIGS efficacy requires genetic approaches. T-DNA insertion mutants (e.g., dcl2-1, dcl4-2e in Arabidopsis) or RNAi lines targeting specific pathway components enable functional dissection [17]. Genotyping protocols for homozygous mutants typically combine gene-specific primers with T-DNA border primers, followed by sequencing confirmation when necessary [17]. Comparative analysis of VIGS efficiency in wild-type versus mutant backgrounds reveals component-specific contributions, as demonstrated by the complete loss of RSV resistance in dcl2 mutant CP transgenic lines despite intact DCL4 function [17]. For species not amenable to stable transformation, virus-based overexpression of viral suppressors of RNA silencing (VSRs) like P19 or HC-Pro can temporarily inhibit specific pathway components and assess their necessity [3].
The coordinated activities of DCL enzymes, RDRP, AGO proteins, and RISC complexes establish the molecular foundation for Virus-Induced Gene Silencing technology, enabling efficient, sequence-specific suppression of target gene expression in plants [3] [1]. Continued research into the structural nuances and regulatory mechanisms governing these core components will further refine VIGS efficacy and expand its applications in functional genomics and crop improvement. Emerging evidence of VIGS-induced epigenetic modifications that can be transmitted transgenerationally presents particularly promising avenues for plant breeding, as DNA methylation established through RNA-directed DNA methylation (RdDM) pathways can create stable, heritable phenotypes [1]. The integration of VIGS with multi-omics technologies and advanced genome-editing platforms will undoubtedly accelerate the discovery and validation of agronomically valuable genes, solidifying the role of these core cellular players as indispensable tools in modern plant biotechnology.
Virus-induced gene silencing (VIGS) is widely recognized as an RNA-mediated reverse genetics technology that operates through post-transcriptional gene silencing (PTGS) in the cytoplasm. However, emerging research has revealed that the implications of VIGS extend far beyond the cytoplasmic realm, directly influencing nuclear epigenetic states through RNA-directed DNA methylation (RdDM). This process mediates transcriptional gene silencing (TGS) of endogenous genomic loci, primarily those related to transposons and repetitive elements, establishing stable, heritable epigenetic modifications [1] [25] [26]. The discovery that VIGS can induce heritable epigenetic marks through the RdDM pathway represents a paradigm shift in our understanding of viral vector applications, transforming VIGS from a transient knockdown tool to a system capable of generating stable epigenetic phenotypes for crop improvement and functional genomics [1].
This technical guide examines the molecular machinery connecting VIGS to RdDM, details experimental methodologies for investigating this pathway, and provides key resources for researchers exploring transcriptional silencing mechanisms within the broader context of VIGS-mediated reverse genetics.
Molecular Mechanisms: From Cytoplasmic Silencing to Nuclear Methylation
The VIGS-RdDM Connection
The conventional VIGS process begins in the cytoplasm with the introduction of recombinant viral vectors carrying target gene sequences. The plant's antiviral defense mechanisms process these into small interfering RNAs (siRNAs) that typically direct mRNA degradation via PTGS [1] [3]. However, when the viral vector insert corresponds to promoter sequences rather than coding regions, a distinct pathway is activated. These promoter-targeting siRNAs can traffic to the nucleus and guide de novo DNA methylation through the RdDM machinery, leading to transcriptional repression of the corresponding gene [1].
This connection was definitively demonstrated when TRV:FWAtr infection in Arabidopsis led to transgenerational epigenetic silencing of the FWA promoter sequence. The silenced state persisted over multiple generations, even after the viral vector was no longer detectable, proving that VIGS can establish stable epigenetic modifications [1].
Core RdDM Pathway Components
The RdDM pathway represents a specialized transcriptional silencing system in plants that employs three distinct DNA-dependent RNA polymerases: Pol II, Pol IV, and Pol V [25].
Figure 1: Molecular pathway of VIGS-induced RdDM showing how viral vectors trigger nuclear DNA methylation.
The pathway initiates with RNA Polymerase IV (Pol IV), which produces precursor transcripts that are converted to double-stranded RNA by RNA-DEPENDENT RNA POLYMERASE 2 (RDR2) [25]. These dsRNAs are processed by DICER-LIKE 3 (DCL3) into 24-nucleotide small interfering RNAs (siRNAs) [25]. The 24-nt siRNAs are loaded into ARGONAUTE 4 (AGO4) or AGO6 to form the effector complex [25]. Simultaneously, RNA Polymerase V (Pol V) generates scaffold transcripts that recruit AGO4-siRNA complexes to specific genomic loci [25]. The interaction between AGO4-bound siRNAs and Pol V transcripts recruits DOMAINS REARRANGED METHYLTRANSFERASE 2 (DRM2), which catalyzes de novo DNA methylation at all cytosine contexts (CG, CHG, and CHH) [1] [25].
Key Regulatory Proteins in RdDM
DTF1/SHH1 represents a critical factor determining the specificity of Pol IV recruitment. This protein contains a homeodomain for potential DNA binding and a SAWADEE domain that recognizes specific histone modifications, particularly H3K9me marks [25] [26]. DTF1 function is required for proper DNA methylation and 24-nt siRNA accumulation at RdDM target loci, with mutation leading to a dramatic reduction in 24-nt siRNAs to approximately 28% of wild-type levels [25].
The DDR complex (comprising DRD1, DMS3, and RDM1) facilitates Pol V transcription by maintaining appropriate chromatin structure. RDM1 also physically associates with AGO4 and DRM2, serving as a bridging component within the effector complex [25].
Table 1: Core Components of the RdDM Pathway and Their Functions
| Component | Type | Key Function | Mutant Phenotype |
|---|---|---|---|
| Pol IV | RNA Polymerase | Produces siRNA precursors | >90% reduction in 24-nt siRNAs [25] |
| Pol V | RNA Polymerase | Generates scaffold transcripts | Loss of transcriptional silencing [25] |
| RDR2 | RNA-dependent RNA Polymerase | Synthesizes dsRNA from Pol IV transcripts | Reduced 24-nt siRNA accumulation [25] |
| DCL3 | Dicer-like Enzyme | Processes dsRNA into 24-nt siRNAs | Loss of 24-nt siRNAs [25] |
| AGO4/6 | Argonaute Protein | siRNA binding and effector complex formation | Reduced DNA methylation and TGS [25] |
| DRM2 | DNA Methyltransferase | Catalyzes de novo DNA methylation | Loss of de novo methylation [1] [25] |
| DTF1/SHH1 | Transcription Factor | Recruits Pol IV to target loci | ~70% reduction in 24-nt siRNAs [25] |
| RDM1 | Scaffold Protein | Bridges AGO4 and DRM2 | Defective in RdDM [25] |
Experimental Evidence and Key Findings
VIGS-Induced Transgenerational Silencing
Seminal research by Bond et al. (2015) demonstrated that TRV:FWAtr infection in wild-type Arabidopsis establishes heritable epigenetic silencing of the FWA promoter [1]. This silencing persisted for multiple generations without the continued presence of the viral vector, indicating stable establishment of epigenetic marks. The system required functional Pol V and the canonical RdDM machinery, as mutations in NRPE1 (the largest subunit of Pol V) completely abolished VIGS-induced RdDM [1].
Fei et al. (2021) further established that virus-induced transcriptional gene silencing (ViTGS)-mediated DNA methylation is fully established in parental lines and faithfully transmitted to subsequent generations [1]. Importantly, their research demonstrated that 100% sequence complementarity between target DNA and small RNAs is not obligatory for transgenerational RdDM, expanding the potential targeting flexibility of this system [1].
Epigenetic Engineering for Crop Improvement
The capacity of VIGS-RdDM to generate stable epigenetic modifications has significant implications for crop improvement. VIGS can now be utilized as a high-throughput tool that induces heritable epigenetic modifications in plants through the viral genome by transiently knocking down targeted gene expression [1]. As a result of DNA methylation progression induced by VIGS, researchers are developing new stable genotypes with desired traits in plants [1].
Table 2: Documented Instances of VIGS-Induced Transcriptional Silencing
| Plant Species | Target Gene/Locus | Silencing Efficiency | Inheritance Pattern | Reference |
|---|---|---|---|---|
| Arabidopsis thaliana | FWA promoter | Stable over generations | Transgenerational | Bond et al. 2015 [1] |
| Camellia drupifera | CdCRY1 (pericarp pigmentation) | ~69.8% at early developmental stage | Transient (tissue-specific) | Plant Methods (2025) [27] |
| Camellia drupifera | CdLAC15 (mesocarp pigmentation) | ~90.91% at mid developmental stage | Transient (tissue-specific) | Plant Methods (2025) [27] |
| Glycine max (Soybean) | GmPDS (phytoene desaturase) | 65-95% (systemic) | Transient | Plants (2025) [7] |
| Various species | Endogenous transposons | Varies by target | Often stable | Multiple studies [1] [26] |
Experimental Protocols for VIGS-RdDM Investigation
Vector Construction for Promoter-Targeted VIGS
To investigate VIGS-induced RdDM, researchers must first construct viral vectors containing promoter sequences rather than coding sequences:
Target Selection: Identify 200-500 bp promoter regions with high specificity to avoid off-target effects. For the FWA experiments, researchers targeted a specific segment of the FWA promoter [1].
Sequence Verification: Use tools like the SGN VIGS Tool (https://vigs.solgenomics.net/) to screen for suitable cleavage sites and ensure specificity through homologous family analysis [27].
Vector Assembly: Clone the promoter fragment into appropriate viral vectors (e.g., TRV2, BPMV, or CLCrV) using restriction enzymes (e.g., EcoRI and XhoI) or recombination-based cloning [7] [27].
Transformation: Introduce recombinant plasmids into Agrobacterium tumefaciens strains (e.g., GV3101) for plant infiltration [7] [27].
Plant Inoculation Methods
Effective delivery of VIGS constructs is crucial for successful RdDM induction:
Agrobacterium Preparation:
- Grow Agrobacterium containing TRV1 and TRV2-derived vectors in YEB medium with appropriate antibiotics (kanamycin, rifampicin) [27].
- Resuspend bacterial pellets in induction buffer (10 mM MES, 10 mM MgCl₂, 200 μM acetosyringone) to OD₆₀₀ = 1.5 [7] [28].
- Incubate at room temperature for 3 hours before infiltration [27].
Inoculation Techniques:
- Cotyledon Node Infiltration: For soybean, bisect sterilized seeds and immerse fresh explants in Agrobacterium suspension for 20-30 minutes [7].
- Pericarp Cutting Immersion: For recalcitrant tissues like Camellia drupifera capsules, this method achieved ~93.94% infiltration efficiency [27].
- Traditional Infiltration: Puncture abaxial side of cotyledons with a needle and flood with Agrobacterium suspension using a needleless syringe [28].
Molecular Validation of RdDM
Confirming successful DNA methylation requires multiple complementary approaches:
Bisulfite Sequencing:
- Treat genomic DNA with sodium bisulfite to convert unmethylated cytosines to uracils.
- Amplify target regions by PCR and sequence to determine methylation status at single-base resolution.
- Calculate percentage methylation for each cytosine context (CG, CHG, CHH).
Small RNA Analysis:
- Extract total RNA using TRIzol or commercial kits.
- Enrich for small RNAs (<200 nt) and prepare libraries for high-throughput sequencing.
- Map 24-nt siRNAs to the target promoter region.
Expression Analysis:
- Monitor transcriptional silencing using RT-qPCR with appropriate reference genes.
- Select stable reference genes validated under VIGS conditions (e.g., GhACT7 and GhPP2A1 in cotton) [28].
- Avoid unstable reference genes like GhUBQ7 and GhUBQ14 which show significant variation under VIGS conditions [28].
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for VIGS-RdDM Investigations
| Reagent/Category | Specific Examples | Function/Application | Considerations |
|---|---|---|---|
| Viral Vectors | TRV (Tobacco Rattle Virus), BPMV (Bean Pod Mottle Virus), CLCrV (Cotton Leaf Crumple Virus) | Delivery of target sequences to host plants | TRV has broad host range; BPMV preferred for legumes [7] [3] |
| Agrobacterium Strains | GV3101, LBA4404 | Delivery of viral vectors to plant cells | GV3101 widely used for VIGS; optimized with appropriate vir genes [7] [27] |
| Antibiotics | Kanamycin, Rifampicin, Gentamicin | Selection of transformed Agrobacterium | Concentrations vary (typically 50 μg/mL kanamycin, 25 μg/mL gentamicin) [28] |
| Induction Compounds | Acetosyringone (200 μM), MES buffer | Induction of vir genes for T-DNA transfer | Critical for efficient transformation [7] [28] |
| Reference Genes (Validated) | GhACT7, GhPP2A1 (in cotton) | Accurate normalization of gene expression in RT-qPCR | Avoid unstable references like GhUBQ7 under VIGS [28] |
| DNA Methylation Analysis | Bisulfite Conversion Kits, DRM2 antibodies | Detection and quantification of DNA methylation | Whole-genome or targeted approaches possible |
| Small RNA Analysis | RNA extraction kits, DCL3 antibodies | Study of 24-nt siRNA biogenesis and accumulation | Enrichment for small RNAs improves detection |
The intersection of VIGS technology with the RdDM pathway has transformed our understanding of plant epigenetic regulation and expanded the toolbox for functional genomics. The ability to engineer heritable epigenetic modifications through transient viral infection represents a powerful approach for both basic research and crop improvement. Future directions will likely focus on refining the specificity of epigenetic targeting, enhancing the stability of induced modifications, and expanding the application of VIGS-RdDM to a broader range of crop species. As these methodologies mature, they hold significant promise for developing novel crop varieties with improved agronomic traits through epigenetic engineering rather than genetic modification.
Virus-induced gene silencing (VIGS) has emerged as an indispensable RNA-mediated reverse genetics technology that enables researchers to elucidate gene function by exploiting plants' innate antiviral defense mechanisms [1] [29]. This powerful technique represents a paradigm shift in functional genomics, allowing for rapid characterization of gene functions without the need for stable plant transformation [30]. The conceptual foundation of VIGS traces back to the observed phenomenon of 'recovery from viral infection' in plants, a term first characterized by van Kammen [1] [29]. However, the transformational moment in VIGS history occurred in 1995 with the landmark development of the first VIGS vector based on Tobacco mosaic virus (TMV) by Kumagai and colleagues [1] [5] [29]. This pioneering work not only established the methodological framework for VIGS but also demonstrated the profound potential of viral vectors as functional genomics tools, ultimately paving the way for modern crop improvement strategies and epigenetic studies [1] [11].
The Foundational Breakthrough: Kumagai et al. (1995)
Experimental Design and Methodology
The seminal 1995 experiment conducted by Kumagai et al. represented a paradigm shift in plant functional genomics methodology. The researchers engineered a recombinant TMV vector to carry a fragment of the phytoene desaturase (PDS) gene from Nicotiana benthamiana (NbPDS) [1] [5]. The experimental workflow involved several critical steps that established the standard protocol for VIGS approaches:
Vector Construction: A 1.5-kb fragment of the NbPDS gene was cloned into the TMV genome, creating a recombinant viral vector capable of expressing plant gene sequences.
In Vitro Transcription: The modified viral genome was transcribed in vitro to produce infectious RNA transcripts.
Plant Inoculation: These transcripts were mechanically inoculated onto N. benthamiana leaves using standard viral inoculation techniques.
Phenotypic Observation: Successful silencing was confirmed by the appearance of a characteristic albino phenotype in newly emerging leaves approximately 2-3 weeks post-inoculation [1] [31].
The selection of PDS as the target gene was strategic, as its silencing produces an easily scorable photobleaching phenotype due to impaired chlorophyll and carotenoid biosynthesis, providing clear visual confirmation of successful gene silencing [5].
Key Findings and Immediate Impact
The TMV-based VIGS system developed by Kumagai et al. achieved several groundbreaking outcomes that would shape the future of plant reverse genetics. Most significantly, the research demonstrated that a recombinant virus could successfully silence endogenous plant genes through sequence-specific degradation of target mRNAs [1]. The appearance of systemic albino phenotypes in inoculated plants provided irrefutable evidence that the silencing signal could travel throughout the plant, affecting tissues far from the initial infection site [1] [5]. This experiment also established that VIGS could achieve substantial downregulation of gene expression without complete knockout, making it possible to study essential genes that might be lethal if completely silenced [30]. The success of this TMV-based approach validated the core concept that plants' antiviral RNA silencing mechanisms could be harnessed as potent tools for functional genomics.
Table 1: Key Characteristics of the First TMV-Based VIGS Vector
| Parameter | Description |
|---|---|
| Viral System | Tobacco Mosaic Virus (TMV) |
| Target Gene | NbPDS (Phytoene desaturase from Nicotiana benthamiana) |
| Insert Size | 1.5-kb fragment of PDS gene |
| Delivery Method | In vitro RNA transcripts mechanically inoculated |
| Time to Phenotype | 2-3 weeks post-inoculation |
| Key Result | Systemic albino phenotype in emerging leaves |
| Historical Significance | First demonstration of engineered virus for endogenous gene silencing |
The Molecular Mechanism of VIGS
Core Silencing Pathway
The molecular machinery underlying VIGS operates through a sophisticated RNA-mediated process that hijacks the plant's innate antiviral defense systems. The process begins when the recombinant viral vector containing the target gene fragment is introduced into the plant cell [1]. During viral replication, double-stranded RNA (dsRNA) molecules are generated either as replication intermediates or through the activity of host RNA-directed RNA polymerase (RDRP) [1] [5]. These dsRNA molecules are recognized by the plant's RNA silencing machinery as foreign, triggering the core VIGS pathway [1]:
Dicer-like enzyme cleaves the dsRNAs into small interfering RNA (siRNA) duplexes approximately 21-24 nucleotides in length [1] [5].
These siRNAs are incorporated into the RNA-induced silencing complex (RISC) where the guide strand directs the complex to complementary mRNA sequences [1] [29].
The catalytic component of RISC, often an Argonaute (AGO) protein, mediates the sequence-specific cleavage and degradation of target mRNAs [1].
The silencing signal amplifies and spreads systemically through the plant, likely mediated by secondary siRNAs produced by host RDRPs, leading to widespread silencing of the target gene [1] [5].
Figure 1: Molecular Mechanism of VIGS. The core pathway showing how recombinant viral vectors trigger sequence-specific gene silencing through the plant's RNA interference machinery.
Epigenetic Extensions: From PTGS to TGS
While the initial VIGS mechanisms primarily operated at the post-transcriptional level (PTGS), recent research has revealed that VIGS can also induce heritable epigenetic modifications through RNA-directed DNA methylation (RdDM) [1] [29]. In this extended pathway, a subset of siRNAs enters the nucleus and guides epigenetic modifiers to target loci with sequence homology [1]. This leads to DNA methylation at cytosine residues in CG, CHG, and CHH contexts, particularly when the viral vector insert corresponds to promoter regions rather than coding sequences [1] [29]. These epigenetic marks can result in transcriptional gene silencing (TGS) and, remarkably, can be stably inherited across generations, creating new stable genotypes with desired traits without altering the underlying DNA sequence [1]. This epigenetic dimension significantly expands the applications of VIGS in plant breeding and functional genomics.
Evolution of VIGS Vector Systems
Beyond TMV: Diversification of Viral Vectors
Following the pioneering TMV vector, researchers have developed numerous VIGS systems based on different viruses, each with distinct advantages and host ranges. The Tobacco rattle virus (TRV) system emerged as a particularly versatile vector that effectively spreads to all plant tissues, including meristems, which many other viruses cannot infect [5]. TRV-based vectors typically employ Agrobacterium-mediated delivery (agroinfiltration) rather than in vitro transcripts, simplifying the inoculation process [5]. Other significant developments include DNA virus-based vectors, such as those derived from geminiviruses like Beet curly top virus (BCTV), which can silence genes in meristem tissue and have exceptionally broad host ranges spanning multiple plant families [32]. More recently, the Cucumber mosaic virus (CMV) has been engineered for VIGS applications in challenging species like banana, achieving 95% infection rates in experimental conditions [33].
Table 2: Evolution of Key VIGS Vector Systems Since the Original TMV Vector
| Vector System | Virus Type | Key Advantages | Notable Applications |
|---|---|---|---|
| TMV [1] | RNA virus | First developed, strong silencing response | N. benthamiana PDS silencing (historical) |
| TRV [5] | RNA virus | Spreads to meristems, mild symptoms, wide host range | Tomato, tobacco, Arabidopsis, functional genomics |
| BCTV [32] | DNA virus (geminivirus) | Broad host range, meristem silencing | Spinach, tomato, non-model systems |
| CMV [33] | RNA virus | Extremely wide natural host range | Banana, monocots, recalcitrant species |
| AMV/TMV/CMV [31] | RNA viruses (30K family) | Small inserts (54bp or less), tunable silencing | N. tabacum, N. benthamiana, partial silencing |
Technical Refinements and Methodological Advances
Vector development has progressed significantly from the original TMV system, with numerous refinements enhancing efficiency, specificity, and applicability. Modern VIGS vectors incorporate sophisticated features such as GATEWAY cloning technology for high-throughput applications and ligation-independent cloning (LIC) systems to simplify vector construction [5]. There has been a concerted effort to develop vectors capable of tunable silencing through the use of smaller inserts (as small as 18-54 bp), enabling researchers to achieve partial rather than complete silencing—particularly valuable for studying essential genes [31]. Delivery methods have also diversified beyond mechanical inoculation of transcripts to include agroinfiltration, biolistic delivery, and specialized techniques for challenging tissues like the pericarp cutting immersion method developed for lignified camellia capsules [27] [32]. These advances have collectively expanded VIGS from a novel phenomenon to a robust, versatile platform for reverse genetics.
The Scientist's Toolkit: Essential Research Reagents and Materials
Table 3: Key Research Reagent Solutions for VIGS Experiments
| Reagent/Material | Function in VIGS | Specific Examples |
|---|---|---|
| Viral Vectors | Carries target gene fragment to trigger silencing | TMV, TRV, BCTV, CMV-based vectors [1] [5] [33] |
| Agrobacterium Strains | Delivery vehicle for DNA-based vectors | Agrobacterium tumefaciens for TRV and CMV systems [5] [33] |
| Enzymes for Molecular Cloning | Vector construction and target gene insertion | Restriction enzymes (AfeI, NheI), DNA ligases, polymerases [31] [33] [27] |
| Selection Antibiotics | Maintain plasmid stability in bacterial cultures | Kanamycin, rifampicin for binary vectors [27] |
| Induction Compounds | Activate Agrobacterium virulence genes | Acetosyringone for agroinfiltration [27] |
| Positive Control Constructs | Validate system functionality | PDS (phytoene desaturase) vectors [31] [5] [33] |
Current Applications and Future Perspectives in Reverse Genetics
The development of that first TMV-based VIGS vector initiated a trajectory of innovation that continues to transform plant functional genomics. Modern VIGS applications extend far beyond initial proof-of-concept studies to address complex biological questions including biotic and abiotic stress responses, metabolic engineering, and epigenetic regulation [1] [30]. The technology has become particularly valuable for characterizing gene function in non-model systems and recalcitrant species where stable transformation remains challenging, including perennial woody plants like Camellia drupifera and important crops like banana [33] [27]. Recent advances demonstrate that VIGS can induce heritable epigenetic modifications that persist across generations, creating new opportunities for crop improvement without permanent genetic alteration [1] [29]. As VIGS technology continues to evolve, it integrates with emerging genome editing approaches, creating synergistic tools that accelerate the pace of discovery in plant biology and agricultural biotechnology.
The historical journey from that first TMV vector to today's sophisticated VIGS systems exemplifies how methodological innovations can transform biological research. What began as a clever exploitation of plant-virus interactions has matured into a powerful reverse genetics platform that continues to expand its capabilities, solidifying its role as an indispensable tool for researchers and drug development professionals working at the intersection of genetics, epigenetics, and agricultural science.
VIGS in Action: Vectors, Delivery Methods, and Breakthrough Applications
Virus-Induced Gene Silencing (VIGS) has emerged as an indispensable RNA-mediated reverse genetics technology for functional genomics in plants. This powerful approach exploits the plant's innate antiviral defense mechanism—specifically, post-transcriptional gene silencing (PTGS)—to achieve targeted knockdown of endogenous genes [1] [34]. The term VIGS was first coined by van Kammen to describe the phenomenon of 'recovery from viral infection' [1], and the technology was pioneered with the development of the first Tobacco mosaic virus (TMV)-based vector by Kumagai et al. in 1995 [1] [5]. The fundamental principle of VIGS involves engineering viral genomes to incorporate fragments of host genes, infecting plants with these recombinant viruses, and leveraging the plant's defense system to silence the corresponding target genes through sequence-specific mRNA degradation [2].
The VIGS process initiates when a recombinant virus containing a fragment of a host gene is introduced into the plant cell. The plant's RNA-dependent RNA polymerase (RDRP) then uses single-stranded viral RNA to produce double-stranded RNA (dsRNA) replication intermediates [35]. These dsRNA molecules are recognized by Dicer-like enzymes (DCL) that cleave them into 21-24 nucleotide small interfering RNAs (siRNAs) [1] [3]. These siRNAs are incorporated into the RNA-induced silencing complex (RISC), which uses them as guides to identify and cleave complementary endogenous mRNA sequences, resulting in gene silencing [1] [35] [5]. A significant advantage of VIGS is its capacity for systemic silencing, as the silencing signal spreads throughout the plant, enabling analysis of gene function in tissues far from the initial infection site [5].
VIGS has revolutionized plant functional genomics by providing a rapid, cost-effective alternative to stable transformation. Unlike traditional genetic approaches that require time-consuming generation of transgenic lines, VIGS can silence target genes within 3-4 weeks, with only partial sequence information needed for effective silencing [5] [34]. This technical advantage makes VIGS particularly valuable for high-throughput functional screening and for studying gene function in plant species that are recalcitrant to stable transformation [7] [27]. The technology has been successfully applied to characterize genes involved in diverse biological processes including disease resistance, abiotic stress tolerance, development, and metabolic pathways [34] [36] [3].
Table 1: Fundamental Characteristics of Major VIGS Vector Systems
| Vector Characteristic | TRV | BPMV | BSMV | DNA Viruses (e.g., CLCrV) |
|---|---|---|---|---|
| Genome Type | Positive-sense single-stranded RNA | Positive-sense single-stranded RNA | Positive-sense single-stranded RNA | Single-stranded DNA |
| Genome Organization | Bipartite (RNA1 & RNA2) | Bipartite | Tripartite (RNAα, RNAβ, RNAγ) | Bipartite (DNA-A & DNA-B) |
| Primary Host Plants | Solanaceae, Arabidopsis, cotton | Soybean, common bean | Barley, wheat, monocots | Cotton, cassava, Arabidopsis |
| Silencing Efficiency | High (65-95%) [7] | High | Moderate to High | Moderate to High |
| Duration of Silencing | 3 weeks to several months | Varies | Varies | Varies |
| Insert Capacity | 300-1500 bp | 300-500 bp | 300-500 bp | 200-500 bp |
| Key Advantages | Broad host range, meristem infiltration, mild symptoms | Soybean-specific, reliable | Effective for monocots, wide cereal host range | Nuclear replication, stable |
Molecular Mechanisms and Vector Systems
RNA Virus-Based VIGS Vectors
Tobacco Rattle Virus (TRV)
Tobacco rattle virus (TRV) has emerged as one of the most versatile and widely used VIGS vectors, particularly for dicotyledonous plants. TRV is a positive-sense single-stranded RNA virus with a bipartite genome consisting of RNA1 and RNA2 [5] [3]. RNA1 encodes two replicase proteins (134 and 194 kDa), a movement protein (MP) that facilitates cell-to-cell movement, and a 16 kDa cysteine-rich protein that functions as a weak suppressor of RNA silencing [5] [3]. RNA2 contains the coat protein (CP) gene and two non-essential genes (29.4K and 32.8K) that can be replaced with multiple cloning sites for inserting target gene fragments [35] [5]. For successful VIGS, both TRV1 and TRV2 components must be delivered to plant cells, typically through Agrobacterium tumefaciens-mediated transformation [5].
The TRV vector system offers several significant advantages that account for its widespread adoption. TRV efficiently infects meristematic tissues, enabling gene silencing in developing flowers and shoot apical meristems—a capability lacking in many other viral vectors [5]. It produces mild viral symptoms in host plants, reducing the potential for confounding phenotypic interpretations [35] [5]. TRV also exhibits a broad host range, successfully silencing genes in numerous plant species across multiple families including Solanaceae (e.g., tomato, tobacco, pepper), Cruciferae (e.g., Arabidopsis), and others [5] [34]. Additionally, TRV-mediated silencing can persist for extended periods, from several weeks to months under optimized conditions [34]. These attributes make TRV particularly valuable for functional genomics studies in model plants and crops such as Nicotiana benthamiana, tomato, pepper, cotton, and Arabidopsis [35] [5] [3].
Barley Stripe Mosaic Virus (BSMV)
Barley stripe mosaic virus (BSMV) is a single-stranded RNA virus with a tripartite genome (RNAα, RNAβ, and RNAγ) that has been developed as a VIGS vector primarily for monocotyledonous plants [34] [2]. RNAα encodes replication proteins, RNAβ contains genes for the coat protein and other movement-associated proteins, and RNAγ encodes a protein involved in viral replication [2]. For VIGS applications, target gene fragments are typically inserted into RNAγ, which is then co-inoculated with RNAα and RNAβ to initiate systemic infection and silencing [2].
BSMV-based vectors have proven particularly valuable for functional genomics studies in cereals and other monocot species where other VIGS systems are ineffective. BSMV has been successfully used to silence genes in barley (Hordeum vulgare), wheat (Triticum aestivum), and the model grass Brachypodium distachyon [34] [2]. For instance, BSMV-VIGS has been employed to investigate the roles of late embryogenesis abundant (LEA) genes such as HVA1 and Dhn6 in drought tolerance in barley, and to validate the function of the high-molecular-weight glutenin subunit gene (HMW-GS 1Bx14) in wheat grain quality [36]. While BSMV has a more limited host range compared to TRV, its efficacy in monocots—which are generally more challenging for VIGS applications—makes it an essential tool for cereal functional genomics.
Bean Pod Mottle Virus (BPMV)
Bean pod mottle virus (BPMV) is a positive-sense single-stranded RNA virus that has been developed as a highly efficient VIGS vector for legumes, particularly soybean (Glycine max) [7] [34]. BPMV has a bipartite genome consisting of RNA1 and RNA2, with RNA1 encoding replication-related proteins and RNA2 containing movement protein and coat protein genes [7]. The BPMV-VIGS system has been extensively optimized for soybean functional genomics, making it the most widely used VIGS system for this important crop species [7].
The BPMV system has been successfully employed to study various aspects of soybean biology, including disease resistance mechanisms. For example, Kandoth et al. utilized BPMV-VIGS to investigate soybean cyst nematode parasitism, while Cooper et al. demonstrated that BPMV-induced silencing of Rpp1 compromised soybean rust immunity [7]. More recently, BPMV has been used to identify the Rsc1-DR gene conferring resistance to soybean mosaic virus strain SC1 (SMV-SC1) and to validate the role of GmBIR1 in enhancing SMV resistance [7]. A significant limitation of traditional BPMV-VIGS is its frequent reliance on particle bombardment for delivery, which can cause leaf phenotypic alterations that interfere with accurate phenotypic evaluation [7]. However, ongoing optimization efforts aim to address this limitation through improved delivery methods.
DNA Virus-Based VIGS Vectors
DNA virus-based vectors, particularly those derived from geminiviruses such as Cotton leaf crumple virus (CLCrV), offer an alternative approach to RNA-based VIGS systems. CLCrV belongs to the genus Begomovirus in the family Geminiviridae and features a bipartite genome composed of two circular single-stranded DNA molecules: DNA-A and DNA-B [35]. DNA-A contains four genes encoding replication-associated protein, transactivator protein, and coat protein, while DNA-B encodes two proteins (BV1 and BC1) involved in nuclear shuttling and intercellular movement of viral DNA [35]. The two components share approximately 200 base pairs of homologous regions known as common regions (CRs) that contain the origin of replication [35].
CLCrV-based VIGS vectors replicate in the nucleus of host cells, producing double-stranded DNA intermediates that serve as templates for further replication and transcription [35]. Unlike RNA viruses that replicate in the cytoplasm, DNA viruses like CLCrV exploit the host's nuclear transcription machinery, which can potentially lead to more stable and persistent silencing in some systems. The CLCrV-VIGS vector was first reported to silence genes through particle bombardment in cotton and has since been applied to functional genomics studies in this important crop species [35]. DNA virus-based vectors are particularly valuable for plant species where RNA virus vectors have limited efficacy, and they offer the advantage of different replication dynamics that may be beneficial for certain applications.
Experimental Protocols and Methodologies
TRV-Mediated VIGS in Soybean
Recent advances have optimized TRV-mediated VIGS for soybean, achieving high silencing efficiency ranging from 65% to 95% [7]. The protocol involves constructing recombinant vectors by amplifying target gene fragments (e.g., GmPDS, GmRpp6907, GmRPT4) from soybean cDNA using specific primers with restriction sites (EcoRI and XhoI) for directional cloning into the pTRV2-GFP vector [7]. The recombinant plasmid is then transformed into Agrobacterium tumefaciens strain GV3101. For inoculation, conventional methods like misting and direct injection show low efficiency due to soybean's thick cuticle and dense trichomes. The optimized protocol uses cotyledon node infection: sterilized soybean seeds are soaked until swollen, longitudinally bisected to obtain half-seed explants, then immersed in Agrobacterium suspensions containing pTRV1 or pTRV2 derivatives for 20-30 minutes [7]. Infection efficiency is monitored via GFP fluorescence, with successful infiltration showing signals in 2-3 cell layers initially, spreading to deeper cells, with >80% of cells exhibiting fluorescence in transverse sections [7]. Silencing phenotypes (e.g., photobleaching for GmPDS) typically appear within 21 days post-inoculation (dpi) [7].
CLCrV-Mediated VIGS in Cotton
For cotton functional genomics, CLCrV-based VIGS provides an effective alternative. The protocol begins with engineering the CLCrV vector by inserting target gene fragments into the DNA-A component [35]. The recombinant CLCrV DNA-A and wild-type DNA-B are then transformed into Agrobacterium tumefaciens strains. Cotton plants at the 2-3 true leaf stage are selected for agroinfiltration. The Agrobacterium cultures carrying DNA-A and DNA-B are mixed in equal ratios (OD₆₀₀ = 1.0) and infiltrated into cotton leaves using a needleless syringe [35]. Successful infection is indicated by the development of leaf crumpling symptoms approximately 10-14 days post-inoculation. Gene silencing efficiency is typically evaluated using visible marker genes such as CLA1 (chloroplast development) or PDS (carotenoid biosynthesis), which produce characteristic bleached phenotypes when silenced [35]. It is important to note that VIGS efficiency may vary between different cotton species, with diploid species (G. arboreum and G. herbaceum) often showing higher silencing efficiency compared to tetraploid species (G. hirsutum and G. barbadense) [35].
BSMV-Mediated VIGS in Monocots
For monocot species, BSMV-mediated VIGS follows a distinct protocol. Target gene fragments (300-500 bp) are cloned into the BSMV γ-vector using appropriate restriction sites or recombination cloning [2]. In vitro transcription is performed to generate infectious RNA transcripts from linearized plasmid DNA containing RNAα, RNAβ, and RNAγ with target inserts. The RNA transcripts are mixed in equal concentrations and applied to carbonundum-dusted leaves of barley, wheat, or other monocot plants through gentle rubbing [34] [2]. Alternatively, for some species, Agrobacterium-mediated delivery of BSMV constructs may be used. Inoculated plants are maintained under controlled environmental conditions (typically 22-25°C with 16-hour light periods) to optimize viral spread and silencing efficiency [34]. Silencing symptoms become apparent 2-3 weeks post-inoculation, and silencing efficiency can be quantified through RT-qPCR analysis of target gene expression levels in comparison to control plants inoculated with empty BSMV vectors [2].
Diagram 1: Molecular mechanism of Virus-Induced Gene Silencing (VIGS). The process involves delivery of recombinant viral vectors, generation of double-stranded RNA intermediates, processing into siRNAs, and ultimately sequence-specific degradation of target mRNAs.
The Scientist's Toolkit: Essential Research Reagents
Table 2: Key Research Reagent Solutions for VIGS Experiments
| Reagent/Material | Function/Purpose | Examples/Specifications |
|---|---|---|
| Viral Vectors | Delivery of target gene fragments to host plants | pTRV1/pTRV2 (TRV), BSMV RNAα/β/γ constructs, pCLCrVA/pCLCrVB (CLCrV), pBPMV constructs |
| Agrobacterium Strains | Delivery of viral vectors to plant cells | GV3101, GV2260, LBA4404 (for Agrobacterium-mediated transformation) |
| Marker Genes | Assessment of silencing efficiency | PDS (photo-bleaching), CLA1 (chloroplast development), GFP (fluorescence), ANS (anthocyanin production) |
| Enzymes for Molecular Cloning | Construction of recombinant VIGS vectors | Restriction enzymes (EcoRI, XhoI), DNA ligases, polymerases for PCR amplification |
| Plant Growth Media | Maintenance of plant materials and Agrobacterium cultures | MS medium, YEB medium (for Agrobacterium), appropriate antibiotics for selection |
| Infiltration Buffers | Facilitating Agrobacterium entry into plant tissues | MMA buffer (10 mM MES, 10 mM MgCl₂, 200 μM acetosyringone, pH 5.6) |
| Detection Reagents | Validation of silencing efficiency | RNA extraction kits, cDNA synthesis kits, qPCR reagents, primers for target genes |
Technical Considerations and Optimization Strategies
Factors Influencing Silencing Efficiency
Several critical factors must be considered when designing VIGS experiments to ensure optimal silencing efficiency. The selection of target gene fragment is paramount—ideal inserts range from 200-500 base pairs with minimal self-complementarity regions to prevent secondary structure formation [34] [2]. The fragment should exhibit high specificity to the target gene while avoiding significant homology to non-target genes to prevent off-target silencing [27]. Insert orientation also affects efficiency, with reverse-oriented inserts often inducing higher silencing efficiency compared to forward-oriented ones [2].
Plant developmental stage at inoculation significantly impacts VIGS success. Younger plants generally show higher silencing efficiency, though optimal stages vary by species [27]. In Camellia drupifera capsules, for instance, optimal VIGS effects for CdCRY1 and CdLAC15 were observed at early (~69.80% efficiency) and mid stages (~90.91% efficiency) of capsule development, respectively [27]. Environmental conditions, particularly temperature, profoundly influence silencing efficiency. Most VIGS systems perform optimally at temperatures between 18-22°C, as higher temperatures can accelerate viral replication but may also enhance plant defense responses that limit silencing persistence [34] [3].
Agrobacterium culture conditions and inoculation methods represent additional optimization points. Optimal optical density (OD₆₀₀) for agroinfiltration typically ranges from 0.3-1.0, with species-specific optima [3]. The addition of acetosyringone (200 μM) to the infiltration medium enhances T-DNA transfer by inducing Agrobacterium virulence genes [27]. For challenging plant species with thick cuticles or dense trichomes (e.g., soybean), alternative inoculation methods such as cotyledon node immersion may significantly improve efficiency over conventional leaf infiltration [7] [27].
Limitations and Counterstrategies
Despite its powerful applications, VIGS technology presents several limitations that researchers must address. Viral symptoms can sometimes mimic or mask silencing phenotypes, particularly when using vectors that cause substantial pathogenicity [34]. This challenge can be mitigated by selecting mild viral vectors like TRV that produce minimal symptoms [5]. Silencing efficiency may vary between plant tissues and developmental stages, with some tissues like meristems potentially showing reduced silencing depending on the vector system [5]. The transient nature of VIGS limits its application to studies requiring short-term gene knockdown, though some systems can maintain silencing for several months under optimized conditions [34].
Off-target silencing represents another significant concern, where sequences with partial homology to the target insert may be unintentionally silenced [2]. This risk can be minimized through careful bioinformatic analysis of insert sequences against the host genome using tools such as the SGN VIGS Tool (https://vigs.solgenomics.net/) [27]. Host range limitations restrict specific VIGS vectors to certain plant families, necessitating vector selection compatible with the target species [2]. Finally, variation in silencing efficiency across plant genotypes highlights the importance of including appropriate controls and potentially optimizing protocols for specific cultivars [35] [7].
The selection of an appropriate VIGS vector—whether TRV, BPMV, BSMV, or DNA viruses like CLCrV—represents a critical decision point in designing effective functional genomics experiments. Each vector system offers distinct advantages and limitations tailored to specific plant families and experimental needs. TRV stands out for its broad host range and efficient meristem infiltration, making it ideal for dicot species. BSMV provides unparalleled utility for monocot functional genomics, while BPMV offers soybean-specific optimization. DNA virus-based vectors like CLCrV present alternative replication dynamics that may benefit certain applications.
Future developments in VIGS technology will likely focus on expanding host ranges, improving silencing persistence, and enhancing specificity. The integration of VIGS with emerging technologies such as CRISPR-based systems and multi-omics approaches holds promise for comprehensive functional analysis. Furthermore, the development of more sophisticated vectors with inducible or tissue-specific silencing capabilities will expand experimental possibilities. As VIGS continues to evolve, it will remain an indispensable tool in plant reverse genetics, enabling researchers to rapidly characterize gene function and accelerate crop improvement programs.
Within the framework of Virus-Induced Gene Silencing (VIGS) and broader RNA-mediated reverse genetics research, the ability to deliver genetic material into plant cells is a foundational step. Agroinfiltration, a technique utilizing Agrobacterium tumefaciens to transfer DNA into plants, is a cornerstone method for transient expression and silencing studies. While model plants like Nicotiana benthamiana are routinely used, applying these techniques to a wider range of species—including recalcitrant crops and specific tissues like woody capsules—demands a versatile toolkit of delivery methods. This guide details three core delivery techniques: syringe infiltration, vacuum infiltration, and cutting-immersion, providing a quantitative and procedural resource for researchers and drug development professionals aiming to implement or optimize these protocols in their functional genomics workflows [3] [27].
Core Agroinfiltration Delivery Techniques: A Comparative Analysis
The choice of delivery method significantly impacts the efficiency and scope of agroinfiltration experiments. The following table summarizes the key characteristics, advantages, and limitations of the three primary techniques.
Table 1: Comparison of Core Agroinfiltration Delivery Techniques
| Technique | Best Suited For | Key Advantage | Primary Limitation | Typical Efficiency |
|---|---|---|---|---|
| Syringe Infiltration | Individual leaves of model plants (e.g., N. benthamiana, Fragaria vesca); localized transformation [37]. | Simplicity; no specialized equipment needed; allows for targeted tissue infiltration. | Labor-intensive for high-throughput studies; not suitable for entire seedlings or delicate tissues. | High in susceptible leaf tissues (e.g., GFP expression confirmed in >44% of transformable F. vesca cells) [37] [38]. |
| Vacuum Infiltration | High-throughput applications; entire seedlings or small plants; uniform tissue saturation. | Excellent for scalable applications like molecular farming; ensures widespread agent distribution. | Requires specialized vacuum equipment; can be stressful for some plant tissues. | Very high (Yields of ~5 g recombinant protein/kg plant biomass reported in N. benthamiana) [39]. |
| Cutting-Immersion | Recalcitrant tissues (e.g., woody capsules, cotyledon nodes); plant species with dense trichomes or thick cuticles [7] [27]. | Overcomes physical barriers to infiltration; effective for hard-to-transform species. | Involves creating a wound, which may introduce stress or contamination risks. | Up to 95% in soybean cotyledon nodes [7] and ~94% in Camellia drupifera capsules [27]. |
Detailed Experimental Protocols
Syringe Infiltration for Leaf Tissue
This protocol, adapted for wild strawberry (Fragaria vesca), is ideal for functional gene analysis in leaf tissue and can be applied to other model plants [37].
Key Research Reagents:
- Agrobacterium Strain: EHA105 demonstrated highest efficiency in F. vesca; GV3101 is also commonly used [37] [7].
- Vector: Binary vector (e.g., pBI121) with gene of interest and reporter (e.g., GFP, GUS) [37].
- Induction Medium: Liquid YEB or LB medium with appropriate antibiotics (e.g., Kanamycin, Rifampicin) and acetosyringone (200 µM), a phenolic compound that induces Agrobacterium's virulence genes [27].
- Infiltration Buffer: 10 mM MgCl₂, 10 mM MES (pH 5.6), and 200 µM acetosyringone.
Methodology:
- Agrobacterium Preparation: Transform the binary vector into the chosen Agrobacterium strain. Inoculate a single colony into induction medium and culture at 28°C with shaking (200-240 rpm) for ~24 hours until OD₆₀₀ reaches 0.9-1.0 [27].
- Cell Harvest and Resuspension: Pellet bacterial cells by centrifugation (5000 rpm for 15 minutes). Resuspend the pellet in infiltration buffer to a final OD₆₀₀ of 0.5-2.0. Let the suspension rest at room temperature for 1-3 hours [37].
- Infiltration: Use a needleless syringe to press the abaxial (lower) side of a fully expanded leaf. Gently inject the bacterial suspension, causing a water-soaked area. For optimal coverage in F. vesca, infiltrate at multiple spots on a single leaf [37].
- Post-Infiltration Handling: Maintain infiltrated plants in high-humidity conditions for 2-3 days. Analyze transient gene expression, typically 2-5 days post-infiltration (dpi) [37].
Cutting-Immersion for Recalcitrant Tissues
This method is optimized for soybean cotyledon nodes and tea oil camellia capsules, proving highly effective for tissues that are difficult to infiltrate [7] [27].
Key Research Reagents:
- Agrobacterium Strain: GV3101 [7].
- VIGS Vector: A two-component system like pTRV1 and pTRV2 is standard for VIGS. pTRV2 carries the target gene insert [7] [3].
- Sterilization Agents: Sodium hypochlorite, ethanol.
- Tissue Culture Media: Solid and liquid MS media for explant recovery and co-cultivation.
Methodology:
- Explant Preparation: For soybean, surface-sterilize seeds and germinate on sterile media. Prepare "half-seed explants" by bisecting swollen sterilized soybeans longitudinally through the cotyledon node [7]. For camellia capsules, create wounds on the pericarp by cutting [27].
- Agrobacterium Preparation: Culture Agrobacterium harboring pTRV1 and pTRV2 constructs as in the syringe protocol. Mix the two cultures in a 1:1 ratio before immersion [7].
- Immersion and Co-cultivation: Immerse the fresh explants or cut capsules in the Agrobacterium suspension for 20-30 minutes with gentle agitation [7]. After immersion, blot the explants dry and transfer them to co-cultivation media for 2-3 days in the dark.
- Transplant and Analysis: Transfer explants to regeneration or rooting media. Silencing phenotypes, such as photobleaching in GmPDS-silenced soybean, can be observed systemically from 14-21 dpi [7].
Technical Specifications and Optimization Parameters
Successful agroinfiltration relies on optimizing key parameters. The following table consolidates critical quantitative data and conditions from recent studies.
Table 2: Technical Specifications and Optimized Parameters for Agroinfiltration
| Parameter | Optimized Condition / Finding | Experimental Context / Impact |
|---|---|---|
| Agrobacterium Density (OD₆₀₀) | 0.5 - 2.0 | A density-dependent antagonism was observed in N. benthamiana; higher total OD can reduce transformation efficiency per strain, suggesting the need for optimization when mixing strains [38]. |
| Co-cultivation Time | 2 - 3 days | Standard period for T-DNA transfer post-infiltration before removing or counter-selecting against Agrobacterium [7] [27]. |
| Acetosyringone Concentration | 200 µM | Critical for inducing the Agrobacterium vir genes; used in both pre-induction and infiltration buffers [27]. |
| Silencing Onset & Duration | Phenotypes visible from 14-21 dpi | Systemic silencing in soybean and camellia using TRV-based VIGS, with effects lasting several weeks [7] [27]. |
| Total Culture OD Effect | Inverse relationship with efficiency | In N. benthamiana, increasing the total OD of a mixed culture from 0.05 to 3.0 reduced the transformation efficiency constant (α) from ~100 to ~6, highlighting significant inter-strain antagonism [38]. |
Integration with VIGS Workflow and Molecular Mechanisms
Agroinfiltration delivery techniques are the critical first step in the VIGS pipeline, enabling the introduction of viral vectors that trigger the plant's RNA interference (RNAi) machinery. The diagram below illustrates the complete workflow from delivery to observable phenotypic change.
Diagram 1: From Agroinfiltration to VIGS Phenotype. This workflow shows how the three delivery techniques introduce Agrobacterium carrying VIGS vectors into plant cells, initiating an RNAi cascade that leads to gene silencing and a measurable phenotype.
The molecular mechanism of VIGS begins after the T-DNA from Agrobacterium is integrated into the plant cell nucleus and viral replication occurs. The resulting double-stranded RNA (dsRNA) is recognized and processed by the plant's defense machinery.
Diagram 2: Molecular Mechanism of RNAi in VIGS. The core Post-Transcriptional Gene Silencing (PTGS) pathway is triggered by viral dsRNA, leading to mRNA cleavage. A secondary pathway can lead to RNA-directed DNA methylation (RdDM) and heritable epigenetic modifications [1].
Essential Research Reagent Solutions
A successful agroinfiltration experiment depends on carefully selected biological and chemical reagents. The following table itemizes key solutions and their functions.
Table 3: Key Research Reagent Solutions for Agroinfiltration and VIGS
| Reagent / Material | Function / Purpose | Examples & Notes |
|---|---|---|
| Agrobacterium Strains | Delivery vehicle for T-DNA transfer. | EHA105: High virulence, often superior efficiency [37]. GV3101: Widely used for VIGS in solanaceous plants and soybean [7]. |
| Viral Vectors | To carry and replicate the target gene insert systemically in the plant. | Tobacco Rattle Virus (TRV): Most widely used VIGS vector due to mild symptoms and broad host range [7] [3]. pTRV1 & pTRV2: The bipartite vector system for TRV [3]. |
| Induction Agents | Activate Agrobacterium vir genes essential for T-DNA transfer. | Acetosyringone: A phenolic compound, used at 150-200 µM in infiltration buffers [27]. |
| Reporter Genes | Visual markers for rapid assessment of transformation/silencing efficiency. | GFP (Green Fluorescent Protein): Confirmed via fluorescence microscopy [37]. GUS (β-glucuronidase): Detected via histochemical staining [37]. PDS (Phytoene desaturase): Silencing causes photobleaching, a visual marker for VIGS efficiency [7]. |
| Infiltration Buffer | Maintains Agrobacterium viability and promotes T-DNA transfer. | Typically contains MgCl₂ (for osmotic balance) and MES buffer (to maintain acidic pH optimal for virulence induction) [37]. |
Virus-induced gene silencing (VIGS) is an RNA-mediated reverse genetics technology that has evolved into an indispensable approach for analyzing gene function in plants [1]. This powerful technique leverages the plant's innate post-transcriptional gene silencing (PTGS) machinery, which naturally functions as an antiviral defense mechanism [3]. When plants encounter viral infections, they recognize and process viral RNA into small interfering RNAs (siRNAs) that guide the sequence-specific degradation of complementary RNA sequences. VIGS exploits this pathway by engineering viral vectors to carry fragments of plant genes, thereby triggering silencing of corresponding endogenous genes [34].
The application of VIGS for high-throughput functional screening represents a significant advancement over traditional genetic approaches. Conventional methods such as T-DNA insertion mutagenesis, chemical mutagenesis, and stable RNAi transformation are often labor-intensive, time-consuming, and limited by gene redundancy [40] [41]. In contrast, VIGS enables transient gene knockdown without the need for stable transformation, allowing for rapid functional characterization of genes within weeks rather than months or years [40] [34]. This efficiency makes VIGS particularly valuable for screening genes involved in complex traits such as biotic and abiotic stress resistance, which are typically governed by multiple genetic factors [34] [41].
The versatility of VIGS has been demonstrated across numerous plant species, including model plants like Nicotiana benthamiana and important crops such as tomato, potato, pepper, cassava, and various monocots [42] [3] [34]. With approximately 35 different VIGS vectors currently available, researchers can select optimal systems for their specific plant species and research questions [40]. The development of VIGS-based forward genetics screening approaches further expands its utility, enabling identification of novel genes involved in critical biological processes without prior knowledge of gene sequence or function [43].
Molecular Mechanisms of VIGS
Fundamental Silencing Pathways
The molecular mechanism of VIGS begins with the delivery of a recombinant virus containing a fragment of a plant gene of interest. This process initiates a sophisticated cellular defense response that ultimately leads to sequence-specific degradation of target mRNAs [1] [34]. The viral vector is introduced into plant cells typically through Agrobacterium-mediated transformation or in vitro transcribed RNA inoculation, after which the virus replicates and spreads systemically throughout the plant [34].
As the recombinant virus replicates, it produces double-stranded RNA (dsRNA) intermediates through the activity of viral or plant-encoded RNA-dependent RNA polymerase (RdRP) [1] [34]. These dsRNA molecules are recognized by plant Dicer-like (DCL) enzymes that process them into small interfering RNAs (siRNAs) of 21-24 nucleotides in length [1]. The siRNAs are then incorporated into the RNA-induced silencing complex (RISC), where they serve as guides for identifying complementary mRNA sequences [1] [34]. Once identified, these target mRNAs are cleaved and degraded, resulting in post-transcriptional gene silencing [1].
The silencing signal amplifies and spreads throughout the plant through the action of host RDRP, which uses the primary siRNA as a template to synthesize secondary dsRNA, leading to the production of secondary siRNAs [1]. This amplification mechanism enhances the silencing effect and contributes to its systemic nature. In some cases, the silencing process can also lead to epigenetic modifications through RNA-directed DNA methylation (RdDM), resulting in transcriptional gene silencing that can sometimes be heritable across generations [1].
Key Molecular Components
The following table summarizes the core molecular components involved in the VIGS pathway:
Table 1: Key Molecular Components in the VIGS Pathway
| Component | Function in VIGS | Characteristics |
|---|---|---|
| Dicer-like (DCL) enzymes | Processes dsRNA into siRNAs | DCL2, DCL3, and DCL4 produce 22-nt, 24-nt, and 21-nt siRNAs, respectively [1] |
| RNA-dependent RNA polymerase (RDRP) | Amplifies silencing by synthesizing secondary dsRNA | Enhances silencing intensity and systemic spread [1] [34] |
| Argonaute (AGO) proteins | Core component of RISC that uses siRNA to identify target RNAs | Different AGO proteins have specialized functions [1] [44] |
| Small interfering RNAs (siRNAs) | Guide sequence-specific recognition and cleavage of target mRNAs | 21-24 nucleotides in length; provide specificity [1] [34] |
Figure 1: Molecular Mechanism of Virus-Induced Gene Silencing (VIGS)
VIGS Toolbox for High-Throughput Screening
Viral Vector Systems
The effectiveness of high-throughput VIGS screening depends significantly on selecting appropriate viral vectors. Different vector systems offer distinct advantages and limitations based on their host range, silencing efficiency, symptom severity, and capacity for foreign DNA insertion [3]. The Tobacco rattle virus (TRV)-based system is currently the most widely used VIGS vector due to its broad host range, efficient systemic movement, mild symptom development, and ability to target meristematic tissues [40] [3] [34].
TRV is a bipartite virus requiring two components: TRV1 encodes replicase and movement proteins, while TRV2 contains the coat protein gene and serves as the insertion site for plant gene fragments [3] [34]. This system has been successfully deployed in numerous Solanaceae species, including Nicotiana benthamiana, tomato, and pepper, as well as in certain monocot species [34]. Other viral vectors include Bean pod mottle virus (BPMV) for legumes, Barley stripe mosaic virus (BSMV) for monocots, and Apple latent spherical virus (ALSV) with a broad host range that includes dicots and monocots [34].
Recent advancements in vector design have addressed previous limitations. For instance, the development of Turnip crinkle virus (TCV)-based vectors like CPB1B enables simultaneous silencing of two different genes with a visualizable silencing indicator, facilitating more complex genetic studies [44]. Similarly, the optimization of Citrus yellow vein-associated virus (CY1)-based vectors through strategic insertion of hairpin sequences with specific thermodynamic properties has improved insert stability, making VIGS more reliable for long-term studies in perennial species [45].
Research Reagent Solutions
Successful implementation of high-throughput VIGS screening requires access to specialized biological materials and reagents. The following table outlines essential components of the VIGS toolkit:
Table 2: Essential Research Reagents for VIGS Screening
| Reagent/Resource | Function/Application | Examples/Specifications |
|---|---|---|
| VIGS Vectors | Delivery of target gene fragments into plant cells | TRV-based (most common), BSMV (monocots), ALSV (broad host range) [40] [3] [34] |
| Agrobacterium Strains | Delivery of viral vectors into plant tissues | GV2260, GV3101 with appropriate virulence [40] [42] |
| cDNA Libraries | Source of gene fragments for forward genetics screening | Normalized libraries from stress-treated tissues [42] [43] |
| VIGS Phenomics Databases | Access to pre-existing silencing phenotype data | VPGD (VIGS Phenomics and Functional Genomics Database) [42] |
| Marker Gene Clones | Silencing controls to assess system efficiency | PDS (photobleaching), ChlH (yellowing), GFP (no visible phenotype) [40] [44] |
The development of VIGS phenomics databases represents a particularly valuable resource for the research community. The VIGS Phenomics and Functional Genomics Database (VPGD) compiles phenotype information from silencing of over 4,000 N. benthamiana genes, with approximately 1,300 genes showing visible phenotypes [42]. This database includes built-in BLAST search functionality, allowing researchers to identify potential phenotypes for genes of interest without performing experiments, thereby streamlining the screening process.
High-Throughput Screening Methodologies
Forward Genetic Screening Approaches
VIGS-based forward genetics screening enables identification of genes involved in specific biological processes without prior knowledge of gene sequence or function. This approach involves silencing random genes from a cDNA library and screening for plants with altered phenotypes or stress responses [43]. A high-throughput methodology has been developed for screening genes involved in nonhost resistance against bacterial pathogens [43].
The forward screening protocol begins with constructing a cDNA library from plants exposed to various biotic and abiotic elicitors, then cloning these fragments into TRV2 vectors [43]. Individual Agrobacterium colonies, each containing a unique TRV2-cDNA construct, are arrayed in 96-well plates and used to inoculate plants along with TRV1 Agrobacterium cultures. This systematic approach allows researchers to screen approximately 100 cDNA clones in 2-3 weeks, with the potential to scale up for larger screens [43].
A key innovation in high-throughput forward screening is the use of GFPuv-expressing pathogens to rapidly identify susceptible plants. Instead of relying on time-consuming bacterial growth assays, researchers can visually assess pathogen growth under UV light as green fluorescent colonies on silenced leaves [43]. This method significantly accelerates the screening process and enhances throughput. Candidate genes identified in primary screens are subjected to secondary validation through individual silencing experiments and conventional pathogen growth assays to confirm their role in stress resistance [43].
Reverse Genetic Screening Approaches
Reverse genetics screening using VIGS involves targeted silencing of candidate genes selected based on prior knowledge, such as transcriptome data from stress-treated plants. This approach is particularly valuable for functional validation of genes identified through omics studies [40] [34]. A high-throughput reverse genetics platform has been developed that combines VIGS with leaf-disk-based stress assays [40].
In this methodology, genes are first silenced in whole plants, then leaf disks are excised and subjected to various abiotic stresses, including drought, salinity, temperature extremes, and oxidative stress [40]. The use of leaf disks rather than whole plants offers several advantages: it enables multiple stress treatments on tissue from the same silenced plant, reduces space requirements, and allows for precise control of stress conditions. Research has confirmed that VIGS progression continues in excised leaf disks for more than six weeks, making this approach reliable for prolonged stress treatments [40].
This reverse screening approach has been successfully applied to characterize 28 different N. benthamiana genes involved in various stress responses, identifying several genes with roles in multi-stress tolerance [40]. The methodology has been validated for studying thermotolerance genes, expanding the application of VIGS to previously challenging stress conditions [40].
Specialized Screening Protocols
Cassava Mosaic Disease Resistance Screening
A specialized VIGS-based rapid screening system has been developed for assessing resistance to cassava mosaic disease (CMD) caused by geminiviruses [46]. This innovative approach uses a VIGS vector derived from East African cassava mosaic virus (EACMV-K201) carrying a fragment of the cassava SPINDLY (MeSPY) gene [46].
When inoculated into CMD-susceptible cassava cultivars, this construct triggers severe shoot-tip necrosis followed by plant death within 2-4 weeks, while CMD-resistant cultivars remain unaffected [46]. The differential response correlates with higher virus titers and reduced MeSPY expression in susceptible plants. This system reduces the screening time from the conventional 12-18 months to just 2-4 weeks, dramatically increasing throughput for CMD resistance breeding programs [46].
Simultaneous Multi-Gene Silencing
For studying complex biological processes involving multiple genes, a novel TCV-based VIGS vector (CPB1B) has been developed that enables simultaneous silencing of two different genes [44]. This vector incorporates a visual silencing marker (a fragment of the PDS gene causing photobleaching) alongside the target gene fragment, allowing researchers to assess silencing penetrance before evaluating the phenotype of interest [44].
The optimal insert size for this system is approximately 100 nucleotides, and it has been successfully used to validate the roles of DCL4 and AGO2 in antiviral defense [44]. This approach is particularly valuable for studying genetic interactions and functional redundancy in stress response pathways.
Experimental Design and Workflow
Standard VIGS Protocol for High-Throughput Screening
The following diagram illustrates the comprehensive workflow for high-throughput VIGS screening:
Figure 2: High-Throughput VIGS Screening Workflow
Critical Experimental Parameters
Successful implementation of high-throughput VIGS screening requires optimization of several key parameters:
Insert Design: Gene fragments of 300-500 bp typically provide optimal silencing efficiency [34]. These fragments should be designed to minimize off-target effects using specialized algorithms and should avoid regions of high sequence similarity with unrelated genes [34].
Agroinfiltration Conditions: The optical density (OD₆₀₀) of Agrobacterium cultures used for infiltration significantly affects silencing efficiency. Typical OD₆₀₀ values range from 0.3 to 1.0, with optimal conditions varying by plant species and vector system [3].
Plant Growth Stage: Plants are most amenable to VIGS at specific developmental stages, typically 3-4 weeks post-germination for most species [40] [42]. Younger plants generally show more efficient silencing, though this must be balanced with the need for sufficient tissue for subsequent stress assays.
Environmental Conditions: Temperature, humidity, and light intensity profoundly influence both viral spread and silencing efficiency. Most VIGS protocols recommend temperatures of 20-22°C during the day and 18-20°C at night, with 60-70% relative humidity and 14-16 hour photoperiods [42] [3].
Stress Application Timing: Stress treatments should be applied when silencing is maximal, typically 2-3 weeks post-inoculation [40]. The duration and intensity of stress must be carefully calibrated to reveal phenotypic differences without causing universal damage.
Applications in Stress Resistance Research
Biotic Stress Resistance Screening
VIGS has emerged as a powerful tool for dissecting the genetic basis of plant immunity against diverse pathogens. In nonhost resistance screening, VIGS has identified numerous genes required for broad-spectrum disease resistance against non-adapted pathogens [43]. This approach has revealed the importance of EDS1, RAR1, and NPR1 in basal defense mechanisms [40].
For specific host-pathogen systems, VIGS enables functional characterization of resistance (R) genes and components of defense signaling pathways. In cassava, a specialized VIGS system has been developed for rapid assessment of resistance to cassava mosaic disease [46]. This approach uses the endogenous MeSPY gene as a silencing target whose effects differ between resistant and susceptible genotypes, enabling rapid discrimination within 2-4 weeks compared to conventional screening that requires 12-18 months [46].
VIGS has also been widely employed to study resistance against fungal pathogens, oomycetes, and insect herbivores. The ability to silence multiple genes simultaneously using advanced vectors like CPB1B facilitates investigation of functional redundancy and pathway interactions in plant immune networks [44].
Abiotic Stress Resistance Screening
The application of VIGS for abiotic stress research has expanded significantly, enabling functional characterization of genes involved in drought, salinity, temperature extremes, and oxidative stress tolerance [40] [34] [41]. The development of leaf-disk-based screening methodologies has been particularly instrumental in advancing high-throughput abiotic stress research [40].
This approach has been used to characterize the roles of numerous abiotic stress-responsive genes, including:
- Drought tolerance: NbP5CS1 (proline biosynthesis), NbCBL1 (calcium signaling), NbMYC2 (transcription factor) [40]
- Salinity tolerance: NbSOS1 (ion homeostasis), NtRAR1 (signaling component), NtNPR1 (salicylic acid signaling) [40]
- Thermotolerance: NbHSP101 (heat shock protein), NbWRKY2 (transcription factor), NbETR1 (ethylene signaling) [40]
- Oxidative stress tolerance: NbRBX1 (redox regulation), NbCTR1 (signaling component), NbAPX3 (antioxidant enzyme) [40]
The versatility of VIGS is further demonstrated by its application in studying nutrient deficiency stress, hypoxia, and UV stress responses [34]. The ability to study multiple stress conditions using leaf disks from the same silenced plant enables researchers to identify genes with specific versus broad functions in stress adaptation [40].
Data Analysis and Validation
Phenotypic Scoring Systems
Robust phenotypic assessment is crucial for successful VIGS screening. For biotic stress responses, standardized scoring systems have been developed to quantify disease symptoms and pathogen growth [43] [46]. The cassava mosaic disease scoring system, for example, uses a 0-5 scale where 0 represents no symptoms and 5 represents severe mosaic distortion and stunting [46].
For abiotic stress screening, phenotypic assessment may include measurements of photosynthetic efficiency, ion leakage (membrane integrity), chlorophyll content, oxidative damage markers, and growth parameters [40] [41]. The leaf-disk-based system enables precise quantification of these parameters under controlled laboratory conditions [40].
High-throughput phenotyping approaches include the use of GFP-labeled pathogens for rapid visual assessment of susceptibility [43] and automated image analysis for quantifying stress-induced morphological changes. These methods significantly enhance screening throughput while maintaining reliability.
Molecular Validation
Phenotypic observations from VIGS screens must be corroborated with molecular evidence to confirm successful silencing of target genes and establish causative relationships. Essential validation steps include:
- Transcript quantification: RT-qPCR analysis to verify reduction in target mRNA levels (typically 70-90% knockdown) [40] [46]
- Viral presence confirmation: PCR amplification of viral coat protein genes to confirm systemic infection [40]
- Protein-level analysis: Western blotting or enzyme activity assays when suitable antibodies or functional assays are available
- Secondary siRNA detection: Northern blotting or high-throughput sequencing to confirm production of siRNAs corresponding to the target gene
For forward genetics screens, identification of the silenced gene requires sequence analysis of the TRV2 insert from susceptible plants [43]. This typically involves PCR amplification from agroinfiltrated leaf tissue followed by Sanger sequencing.
Hit Confirmation
Candidate genes identified in primary screens require validation through independent experiments:
- Independent VIGS: Repeating the silencing experiment with newly generated vectors containing different fragments of the same gene [43]
- Stable transformation: Complementation tests with overexpression constructs or confirmation with stable RNAi lines [41]
- Multiple pathogen/stress testing: Assessing whether susceptibility or sensitivity extends to multiple stressors [40] [43]
- Cross-species validation: Testing whether gene function is conserved across related species using heterologous silencing [41]
This rigorous validation process ensures that phenotypes are reproducible and specific to the targeted gene rather than resulting from off-target effects or experimental artifacts.
Advanced Applications and Future Perspectives
Integration with Emerging Technologies
The power of VIGS-based screening is greatly enhanced when integrated with other functional genomics technologies. The availability of the N. benthamiana genome sequence facilitates genome-wide silencing screens and precise prediction of off-target effects [42]. Combining VIGS with high-throughput phenotyping platforms enables automated, quantitative assessment of complex traits at scale.
Integration with multi-omics approaches (transcriptomics, proteomics, metabolomics) allows for comprehensive characterization of molecular consequences following gene silencing, providing insights into biological pathways and networks [3]. The development of VIGS phenomics databases represents another significant advancement, enabling researchers to leverage existing phenotype data to guide experimental design [42].
Emerging applications include the use of VIGS for functional characterization of non-coding RNAs, investigation of epigenetic regulation through virus-induced transcriptional gene silencing (ViTGS), and analysis of protein complexes through silencing of individual components [1].
VIGS in Crop Improvement Programs
VIGS-based screening has significant potential for accelerating crop improvement programs by rapidly identifying genes that confer desirable agronomic traits. In pepper (Capsicum annuum), VIGS has been used to characterize genes controlling fruit quality (color, biochemical composition, pungency), resistance to biotic stresses (bacteria, oomycetes, insects), and tolerance to abiotic stresses (temperature, salt, osmotic stress) [3].
The ability of VIGS to function in transient assays without stable transformation is particularly valuable for crops with long life cycles or recalcitrant transformation systems, such as cassava and fruit trees [45] [46]. Recent developments in vector design have improved insert stability, making VIGS more reliable for long-term studies in perennial species [45].
Looking forward, VIGS is poised to play an important role in functional characterization of candidate genes identified through genome-wide association studies (GWAS) and genomic selection, bridging the gap between genotype and phenotype in crop breeding programs [3].
Current Limitations and Future Directions
Despite its considerable advantages, VIGS still faces several limitations that require attention. Insert stability remains a challenge for some vector systems, though recent advances in understanding RNA thermodynamic properties have led to improved designs [45]. Variability in silencing efficiency between experiments and between individual plants can complicate interpretation, necessitating appropriate experimental designs with sufficient replication.
Tissue-specific variation in silencing efficiency and inability to silence all gene families equally represent additional challenges. Future developments will likely focus on expanding the range of amenable species, improving silencing consistency, developing inducible systems for silencing essential genes, and creating more sophisticated vectors with tissue-specific expression or the ability to silence multiple genes simultaneously [44].
The ongoing refinement of VIGS technology promises to further enhance its utility for high-throughput functional screening, cementing its position as an essential tool for plant functional genomics and stress resistance research.
Virus-Induced Gene Silencing (VIGS) has emerged as an indispensable RNA-mediated reverse genetics technology for analyzing gene function in plants. This powerful approach utilizes the natural antiviral defense mechanism of plants, known as post-transcriptional gene silencing (PTGS), to achieve systemic suppression of endogenous gene expression [1] [3]. The fundamental principle of VIGS involves engineering viral vectors to carry fragments of plant genes, which when introduced into the plant, trigger a sequence-specific RNA degradation mechanism that silences the corresponding endogenous genes [1] [47]. This technology has revolutionized functional genomics in species that are recalcitrant to stable genetic transformation, including medicinal plants like Catharanthus roseus [3] [48].
The historical development of VIGS dates back to 1995 when Kumagai et al. first used a Tobacco mosaic virus (TMV) vector carrying a fragment of the phytoene desaturase (PDS) gene from Nicotiana benthamiana to induce silencing, resulting in a characteristic photo-bleaching phenotype [3]. Since this pioneering work, VIGS has been adapted for use in a diverse array of plant species, with the toolkit expanding significantly to include vectors based on various viruses such as Tobacco Rattle Virus (TRV), Alfalfa mosaic virus (AMV), and Cotton leaf crumple virus (CLCrV) [3]. To date, VIGS has been successfully applied for functional gene analysis in over 50 plant species, including major crops and medicinal plants, enabling the characterization of hundreds of genes involved in disease resistance, abiotic stress responses, and specialized metabolism [3].
For medicinal plant research, VIGS offers particular advantages because these species often have complex genomes, long generation times, and are frequently recalcitrant to stable transformation [48]. The ability to rapidly silence genes without the need for stable transformation makes VIGS an ideal tool for elucidating biosynthetic pathways of valuable specialized metabolites, which has profound implications for drug development and sustainable production of plant-derived pharmaceuticals [49] [48].
Molecular Mechanisms of VIGS
Core Silencing Machinery
The biological basis of VIGS is the conserved RNA silencing mechanism that plants employ as an antiviral defense system [3]. The process begins when a recombinant viral vector containing a fragment of a plant gene is introduced into the plant cell. Upon viral transcription and replication, double-stranded RNA (dsRNA) molecules are produced in the plant cell cytoplasm [47]. These dsRNA molecules are recognized by the plant's Dicer-like (DCL) enzymes, which cleave them into small interfering RNAs (siRNAs) of approximately 21-24 nucleotides in length [1].
These siRNAs are then incorporated into an RNA-induced silencing complex (RISC), which contains Argonaute (AGO) proteins as catalytic components [1] [3]. The RISC complex uses the siRNAs as guides to specifically identify and cleave complementary viral RNA sequences, thereby suppressing infection. In the case of VIGS, this machinery also targets complementary endogenous mRNA transcripts for degradation, leading to silencing of the plant gene of interest [1] [3]. According to studies, secondary siRNAs produced through the cleavage of dsRNA synthesized by the host RNA-dependent RNA polymerase (RDRP) using the primary siRNA as a template appear to enhance VIGS maintenance and dissemination throughout the plant [1].
Transcriptional Gene Silencing and Epigenetic Modifications
Beyond post-transcriptional silencing, VIGS can also induce heritable epigenetic modifications in plants through RNA-directed DNA methylation (RdDM) [1]. In this mechanism, the AGO complex can interact with target DNA molecules in the nucleus, causing transcriptional repression via DNA methylation at the 5' untranslated region (5'UTR), which results in transcriptional gene silencing (TGS) [1].
This epigenetic aspect of VIGS involves DNA methyltransferases that introduce methyl groups on cytosine residues at CG, CHG, and CHH contexts [1]. These methyl groups can result in heritable gene silencing if they are in proximity to promoter sequences [1]. The VIGS-induced epigenetic modifications can be stably transmitted to subsequent generations, creating new stable genotypes with desired traits without altering the underlying DNA sequence [1]. This aspect of VIGS has significant implications for plant breeding programs and sustainable crop improvement, including the enhancement of medicinal compounds in plants [1].
Table 1: Key Components of the VIGS Machinery
| Component | Function | Role in VIGS Process |
|---|---|---|
| Dicer-like (DCL) enzymes | Cleaves dsRNA into siRNAs | Initiates processing of viral dsRNA into guide molecules |
| Small interfering RNAs (siRNAs) | 21-24 nt guide molecules | Directs sequence-specific recognition of target mRNA |
| Argonaute (AGO) proteins | Catalytic component of RISC | Mediates cleavage or translational inhibition of target mRNA |
| RNA-induced silencing complex (RISC) | Effector complex | Executes mRNA degradation or translational repression |
| RNA-dependent RNA polymerase (RDRP) | Amplifies dsRNA | Produces secondary siRNAs for signal amplification |
| DNA methyltransferases | Adds methyl groups to DNA | Mediates transcriptional gene silencing and epigenetic modifications |
VIGS Applications in Catharanthus roseus
Studying Terpenoid Indole Alkaloid Biosynthesis
Catharanthus roseus (Madagascar periwinkle) is a highly valued medicinal plant that produces nearly 200 terpenoid indole alkaloids (TIAs), including the critically important anti-cancer drugs vinblastine and vincristine [48]. The TIA biosynthetic pathway in C. roseus is exceptionally complex, requiring over 30 enzymes, transport within cellular compartments and among multiple cell types, and competing flux towards variable end-products [50]. This complexity, combined with the difficulty of generating stable transgenic C. roseus plants, makes VIGS an indispensable tool for elucidating this pathway [49] [48].
The application of VIGS in C. roseus has enabled researchers to functionally characterize genes encoding biosynthetic enzymes, transporters, and regulators involved in TIA biosynthesis [48]. For instance, VIGS has been used to investigate the vindoline pathway - a crucial branch of TIA biosynthesis leading to the formation of vindoline, which is a precursor for vinblastine and vincristine [49]. This pathway consists of seven enzymes: T16H2, 16OMT, T3O, T3R, NMT, D4H, and DAT [50]. Unlike the upstream TIA biosynthetic pathway, which is directly regulated by jasmonic acid (JA) signaling transcription factors, the vindoline pathway is not directly regulated by these factors and is instead activated by light, leaf-specific differentiation, and developmental state [50] [51].
Key Findings from VIGS Studies in C. roseus
Several transcription factors regulating TIA biosynthesis have been characterized using VIGS in C. roseus. For example, silencing of CrGATA1, a GATA-type zinc-finger transcription factor that activates the vindoline pathway in response to light, led to downregulation of vindoline pathway genes (T3O, T3R, and DAT) and decreased vindoline contents in cotyledons [48]. Similarly, silencing of CrMYC2, a key regulator in the jasmonate response, followed by elicitation with methyl jasmonate (MeJA), resulted in the downregulation of ORCA2 and ORCA3, which are transcription factors that activate upstream TIA pathway genes [48].
Recent research has also identified a GLK homologue (CrGLK) in C. roseus and used VIGS to characterize its role in regulating TIA biosynthesis [51]. Reducing CrGLK levels via VIGS or the chloroplast retrograde signaling inducer lincomycin increased TIA accumulation and TIA pathway gene expression, suggesting that CrGLK may repress TIA biosynthesis [51]. This finding contributes to our understanding of how plants might coordinate chlorophyll and alkaloid biosynthesis during seedling and leaf development.
Additionally, VIGS has been employed to study the role of DELLA proteins, which are key regulators of gibberellic acid (GA) signaling, in C. roseus [50]. Silencing CrDELLA genes resulted in elongated plants, while silencing CrGID1 genes (which encode GA receptors that promote DELLA degradation) produced dwarfed plants, consistent with their roles in GA-mediated growth [50]. In initial experiments, CrDELLA-silencing significantly decreased vindoline pathway gene expression, while CrGID1-silencing significantly increased vindoline, catharanthine, ajmalicine, and serpentine accumulation, although subsequent experiments found little to no effect, highlighting the complexity of these regulatory networks [50].
Table 2: Key Transcription Factors in C. roseus Studied Using VIGS
| Transcription Factor | Regulatory Role | Effect of Silencing | Reference |
|---|---|---|---|
| CrGATA1 | Activates vindoline pathway in light | Downregulation of vindoline pathway genes and decreased vindoline accumulation | [48] |
| CrMYC2 | Mediates jasmonate response | Downregulation of ORCA2 and ORCA3 after MeJA elicitation | [48] |
| CrGLK | Regulates chloroplast development | Increased TIA accumulation and pathway gene expression | [51] |
| CrGBF1/CrGBF2 | Repressors of TIA biosynthesis | Upregulation of TIA pathway genes when silenced | [48] |
Experimental Protocols for VIGS in C. roseus
Cotyledon-Based VIGS Method
A significant advancement in VIGS for C. roseus is the development of a cotyledon-based VIGS (cotyledon-VIGS) approach, which is significantly faster and more efficient than previously described methods [48]. The protocol involves using five-day-old, etiolated seedlings of C. roseus that are vacuum-infiltrated with Agrobacterium tumefaciens GV3101 cells carrying the TRV vectors [48]. The detailed methodology is as follows:
Seed Germination: C. roseus seeds are germinated in the dark at appropriate temperature conditions. Radicles emerge from the seed coats on the second day, while cotyledons fully emerge on the fifth day [48].
Vector Preparation: The bipartite Tobacco Rattle Virus (TRV) system is used, consisting of TRV1 and TRV2 vectors. The gene fragment of interest (typically 100-500 bp) is cloned into the TRV2 vector [3] [31]. For the marker gene, protoporphyrin IX magnesium chelatase subunit H (ChlH), involved in chlorophyll biosynthesis, can be used as it produces a visible yellow phenotype upon silencing [48].
Agrobacterium Preparation: Agrobacterium tumefaciens GV3101 strains containing TRV1 and TRV2 (with insert) vectors are grown overnight in appropriate media with antibiotics. The cultures are then resuspended in infiltration medium (e.g., 10 mM MgCl₂, 10 mM MES, 200 μM acetosyringone) to an OD₆₀₀ of 1.0 [48].
Vacuum Infiltration: Five-day-old seedlings are submerged in the Agrobacterium suspension and placed in a vacuum chamber. The vacuum is applied for 30 minutes to facilitate infiltration [48].
Post-Infiltration Care: Following infiltration, the sprouts or seedlings are kept in the dark until they are 8-day-old and then exposed to light. A clear yellow phenotype can be observed in cotyledons after silencing ChlH when seedlings are first grown in the dark, followed by 2-3 days of light exposure [48].
This cotyledon-VIGS method has been successfully extended to other valuable medicinal plants, including Glycyrrhiza inflata (licorice) and Artemisia annua (sweet wormwood), indicating its broad applicability [48].
Traditional VIGS Methods
Before the development of cotyledon-VIGS, several inoculation methods were tested for C. roseus, with only the pinch wounding method proving successful [48]. This method involves:
Plant Material: 2-3 week-old C. roseus plants are used [1].
Inoculation: The shoot apical meristem and youngest leaves are pinched with fine forceps dipped in Agrobacterium suspension carrying TRV vectors [1].
Silencing Timeline: Silencing becomes evident approximately 2-3 weeks post-inoculation [1].
This method, while effective, is more time-consuming and less efficient than the cotyledon-VIGS approach, which shows silencing phenotypes in as little as 6 days after infiltration [48].
The Scientist's Toolkit: Essential Research Reagents
Successful implementation of VIGS in medicinal plants requires specific reagents and vectors optimized for the target species. The table below details key research reagent solutions for VIGS experiments in C. roseus and other medicinal plants.
Table 3: Essential Research Reagents for VIGS in Medicinal Plants
| Reagent/Vector | Function | Application in C. roseus | |
|---|---|---|---|
| TRV Vectors (TRV1, TRV2) | Bipartite viral vector system | Most widely used VIGS vector for C. roseus; provides broad host range and efficient silencing | [3] [48] |
| Agrobacterium tumefaciens GV3101 | Delivery of TRV vectors into plant cells | Standard strain for C. roseus VIGS; optimized for vacuum and pinch wounding infiltration | [48] |
| Marker Genes (ChlH, PDS) | Visual indicators of silencing efficiency | ChlH silencing produces yellow cotyledons; PDS silencing causes photobleaching | [48] |
| Infiltration Medium (10 mM MgCl₂, 10 mM MES, 200 μM acetosyringone) | Preparation of Agrobacterium suspension for infiltration | Maintains bacterial viability and enhances T-DNA transfer during vacuum infiltration | [48] |
| Methyl Jasmonate (MeJA) | Elicitor of jasmonate-responsive pathways | Used in combination with VIGS to study JA-responsive transcription factors like CrMYC2 | [48] |
| Lincomycin | Inducer of chloroplast retrograde signaling | Used to validate VIGS results for chloroplast-related genes like CrGLK | [51] |
Signaling Pathways in C. roseus Elucidated via VIGS
VIGS studies in C. roseus have been instrumental in elucidating complex signaling pathways that regulate TIA biosynthesis. The diagrams below illustrate key pathways and regulatory networks uncovered through VIGS experiments.
Light and Jasmonate Signaling Networks
Light and JA Signaling in C. roseus
VIGS Experimental Workflow
VIGS Experimental Workflow
Current Advancements and Future Perspectives
Novel Vector Engineering
Recent advancements in VIGS technology include the development of innovative vector systems that enhance silencing efficiency and expand host range. One promising approach involves modifying 30K family movement proteins (MPs) to create VIGS vectors capable of inducing gene silencing with smaller inserts (54 bp or less) compared to the traditional 100-500 bp fragments [31]. This strategy, demonstrated with alfalfa mosaic virus (AMV), cucumber mosaic virus (CMV), and tobacco mosaic virus (TMV), enables calibration of silencing levels based on insert size and offers new opportunities for partial silencing of essential genes [31].
The 30K family of MP comprises 20 viral genera and over 500 viral species that can infect all agronomically significant plant species, making this strategy potentially applicable to a wide range of relevant hosts, including medicinal plants [31]. This approach also revealed an inverse relationship between viral encapsidation efficiency and gene silencing level, providing important insights for vector optimization [31].
Integration with Other Technologies
The future of VIGS in medicinal plant research lies in its integration with other cutting-edge technologies. VIGS is increasingly being combined with transient gene overexpression to achieve simultaneous up- and down-regulation of desired genes in non-model plants [48]. This approach was demonstrated in C. roseus seedlings that were co-infiltrated with TRV vectors silencing two repressors (CrGBF1 and CrGBF2) while overexpressing an activator (CrMYC2), resulting in significant upregulation of TIA pathway genes [48].
Additionally, the emergence of Spray-Induced Gene Silencing (SIGS) represents another frontier with potential applications in medicinal plants [52]. SIGS involves direct application of dsRNAs or siRNAs onto host plants to silence target genes in pathogens or the plants themselves, offering a non-transgenic alternative to VIGS [52]. While primarily developed for crop protection, SIGS principles could be adapted to manipulate specialized metabolism in medicinal plants.
There is also growing interest in combining VIGS with CRISPR/Cas9 technology through virus-mediated delivery of gene editing components, an approach known as Virus-Induced Gene Editing (VIGE) [47]. This integration could enable precise genome editing in medicinal plants that are recalcitrant to stable transformation, opening new avenues for metabolic engineering of high-value pharmaceutical compounds.
VIGS has established itself as a cornerstone technology for functional genomics in medicinal plants, particularly for species like Catharanthus roseus that produce valuable specialized metabolites but are challenging to transform stably. The continued refinement of VIGS methodologies, including the development of more efficient inoculation protocols like cotyledon-VIGS and novel vector engineering approaches, has significantly expanded our ability to decipher complex metabolic pathways. The integration of VIGS with other technologies such as transient overexpression, SIGS, and CRISPR-based editing promises to further accelerate the pace of discovery in medicinal plant research. As these tools continue to evolve, VIGS will play an increasingly vital role in unlocking the genetic potential of medicinal plants, facilitating the discovery and sustainable production of valuable therapeutic compounds for drug development.
Virus-induced gene silencing (VIGS) has evolved from a functional genomics tool into a powerful technology for inducing heritable epigenetic modifications in plants. As an RNA-mediated reverse genetics technique, VIGS traditionally utilizes the plant's post-transcriptional gene silencing (PTGS) machinery to degrade targeted mRNA sequences, enabling rapid functional analysis of genes. Recent breakthroughs have demonstrated that VIGS can be engineered to achieve stable transcriptional gene silencing (TGS) through the induction of RNA-directed DNA methylation (RdDM), leading to transgenerational inheritance of epigenetic states [1]. This technical advancement positions VIGS as a unique tool for creating stable epigenetic variation without altering the underlying DNA sequence, with significant implications for crop improvement and epigenetic research.
The capacity to induce heritable epigenetic marks through VIGS represents a paradigm shift in plant biotechnology. Unlike conventional VIGS that results in transient knockdown of gene expression, VIGS-induced epigenetic silencing can persist for multiple generations, providing a powerful method for studying transgenerational inheritance and developing novel plant varieties with improved agronomic traits [1]. This technical guide explores the molecular mechanisms, experimental protocols, and applications of VIGS-mediated heritable epigenetics within the broader context of VIGS RNA-mediated reverse genetics technology research.
Molecular Mechanisms of VIGS-Induced Heritable Epigenetic Modifications
From Post-Transcriptional to Transcriptional Gene Silencing
The conventional VIGS mechanism operates primarily at the post-transcriptional level in the cytoplasm. When a recombinant viral vector carrying a fragment of a target gene is introduced into a plant, the plant's antiviral defense machinery is activated. This process involves the recognition of viral double-stranded RNA (dsRNA) by Dicer-like enzymes (DCL), which process them into 21-24 nucleotide small interfering RNAs (siRNAs) [1] [53]. These siRNAs are incorporated into the RNA-induced silencing complex (RISC), which guides the sequence-specific cleavage and degradation of complementary mRNA targets, resulting in PTGS [1] [5].
For heritable epigenetic modifications, VIGS must transition from cytoplasmic PTGS to nuclear transcriptional gene silencing. This occurs when virus-derived siRNAs or secondary siRNAs are translocated to the nucleus and guide the RdDM machinery to homologous DNA sequences [1]. The critical distinction is that for epigenetic silencing, the viral vector insert must correspond to the promoter region of the target gene rather than the coding sequence, directing DNA methylation to regulatory regions and potentially inducing long-term stable silencing [1].
RNA-Directed DNA Methylation and Epigenetic Memory
The establishment of heritable epigenetic marks through VIGS centers on the RdDM pathway, which mediates de novo DNA methylation. As illustrated in Figure 1, this process involves multiple specialized components:
Figure 1: Molecular mechanism of VIGS-induced heritable epigenetic silencing
The core mechanism involves Pol V-dependent scaffold RNAs that serve as binding platforms for AGO4-siRNA complexes, recruiting DNA methyltransferases such as DRM2 to establish de novo methylation at all cytosine contexts (CG, CHG, and CHH) [1] [53]. Once established, this methylation can be maintained across generations through both RNA-independent (involving MET1 and CMT3 methyltransferases) and RNA-dependent maintenance mechanisms (involving canonical Pol IV-RdDM) [1]. The heritability of these epigenetic marks depends on several factors, including the sequence context of the target locus and the activity of maintenance mechanisms in subsequent generations.
Experimental Protocols for VIGS-Induced Epigenetic Silencing
Vector Design and Construction for Epigenetic Silencing
Successful induction of heritable epigenetic modifications requires careful vector design. Unlike conventional VIGS that targets coding sequences, epigenetic VIGS must be directed to promoter regions of target genes. The tobacco rattle virus (TRV)-based vector system remains the most widely used for this application due to its broad host range and ability to target meristematic tissues [3] [5].
Key steps in vector construction:
- Identify target promoter sequence: Select a 200-300 bp fragment from the promoter region of the target gene with high specificity to avoid off-target silencing [54] [27].
- Clone into TRV2 vector: Insert the promoter fragment into the multiple cloning site of the TRV2 vector using appropriate cloning strategies (Gateway, LIC, or traditional restriction digestion) [5].
- Transform Agrobacterium: Introduce both TRV1 (containing replication and movement proteins) and recombinant TRV2 vectors into Agrobacterium tumefaciens strain GV3101 [54].
- Verify construct sequence: Confirm the integrity of the insert and vector components through sequencing before plant inoculation [27].
Critical considerations: The selected promoter fragment should be analyzed for C-residue density, as targets with high percentages of cytosines in CG context show improved RNA-independent maintenance of methylation [1]. Additionally, bioinformatic analysis should ensure the fragment has minimal similarity to other genomic sequences to prevent unintended silencing.
Plant Inoculation and Optimization Parameters
Effective delivery of VIGS constructs is essential for inducing epigenetic modifications. Multiple inoculation methods have been developed for different plant species and tissue types:
Table 1: Comparison of VIGS Inoculation Methods for Epigenetic Applications
| Method | Procedure | Optimal Parameters | Efficiency Range | Best Suited For |
|---|---|---|---|---|
| Leaf Infiltration | Syringe infiltration of Agrobacterium suspension into leaves | OD₆₀₀ = 0.5-1.5; 4-6 leaf stage | 40-70% | Model plants (N. benthamiana, Arabidopsis) [54] [5] |
| Spray Infiltration | Spraying Agrobacterium suspension with surfactant on seedlings | OD₆₀₀ = 1.0-1.5; 5-10 true leaves | 45-75% | High-throughput applications, walnut, pepper [54] [3] |
| Pericarp Cutting Immersion | Immersion of cut fruit tissues in Agrobacterium suspension | OD₆₀₀ = 1.0; early developmental stages | Up to 94% | Recalcitrant fruits (Camellia drupifera) [27] |
| Vacuum Infiltration | Submersion of whole plants under vacuum | OD₆₀₀ = 0.8-1.0; young seedlings | 50-80% | Small Arabidopsis plants, seedlings [5] |
Multiple factors significantly influence silencing efficiency and must be optimized for each species:
- Agrobacterium density: Optimal OD₆₀₀ typically ranges from 0.8 to 1.5, with specific optima for different species (e.g., OD₆₀₀ = 1.1 for walnut) [54].
- Plant developmental stage: Younger tissues generally show higher silencing efficiency, though the optimal stage varies (e.g., 5-10 leaf stage for spray infiltration) [54].
- Temperature: Standard TRV vectors function best at 19-25°C, though California TRV variants maintain efficiency up to 28-30°C [55].
- Fragment length: Inserts of 200-300 bp typically yield optimal silencing, with longer fragments potentially reducing efficiency [54] [27].
Confirmation and Analysis of Epigenetic Modifications
Validating successful epigenetic silencing requires multiple molecular analyses across generations:
Primary generation (T₀) analysis:
- Phenotypic assessment: Document visible phenotypes associated with target gene silencing.
- mRNA expression analysis: Quantify transcript levels using qRT-PCR to confirm knockdown.
- siRNA detection: Verify the presence of target-specific 21-24 nt siRNAs by northern blot or sequencing.
Epigenetic analysis:
- Bisulfite sequencing: Analyze DNA methylation patterns in the target promoter region to confirm RdDM.
- Chromatin immunoprecipitation: Assess histone modifications (H3K9me2, H3K27me3) associated with silent chromatin.
Transgenerational analysis (T₁, T₂ generations):
- Stability of silencing: Monitor phenotype and gene expression in subsequent generations without viral vector.
- Methylation maintenance: Track persistence of DNA methylation patterns.
- Segregation analysis: Determine inheritance patterns of the silent epigenetic state.
Applications and Case Studies
Proof of Concept: FWA Epigenetic Silencing in Arabidopsis
The pioneering demonstration of VIGS-induced heritable epigenetics came from studies on the FWA gene in Arabidopsis. Bond et al. (2015) used TRV vectors carrying fragments from the FWA promoter (TRV:FWAtr) to induce de novo DNA methylation and stable silencing of the FWA gene [1]. This resulted in a heritable late-flowering phenotype that persisted for multiple generations without the presence of the viral vector. Molecular analysis confirmed that the silencing was associated with increased DNA methylation specifically in the FWA promoter region, demonstrating the efficacy of VIGS in establishing stable epigenetic states [1].
This case study established critical parameters for VIGS-induced epigenetics, including the requirement for promoter-targeting sequences and the importance of the Pol V pathway for maintaining silencing. Mutants defective in Pol V failed to establish stable epigenetic silencing, highlighting the essential role of the canonical RdDM pathway in this process [1].
Applications in Crop Species
VIGS-induced epigenetic modifications have been successfully applied in various crop species to improve agronomic traits:
Table 2: Documented Applications of VIGS-Induced Epigenetic Modifications
| Plant Species | Target Gene | Trait Modified | Silencing Efficiency | Inheritance Duration |
|---|---|---|---|---|
| Arabidopsis thaliana | FWA | Flowering time | >70% | At least 2 generations [1] |
| Linaria vulgaris | Lcyc | Flower symmetry | Not specified | Natural epiallele example [1] |
| Juglans regia (walnut) | JrPOR | Chlorophyll synthesis | ~48% | Transient [54] |
| Camellia drupifera | CdCRY1, CdLAC15 | Pericarp pigmentation | 70-91% | Transient [27] |
| Capsicum annuum (pepper) | Multiple genes | Fruit quality, disease resistance | Variable | Transient [3] |
These applications demonstrate the potential of VIGS-mediated epigenetics for crop improvement, particularly for species recalcitrant to stable transformation. The technology enables the creation of novel epialleles with desired traits that can be stabilized through selective breeding.
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagents for VIGS-Induced Epigenetic Studies
| Reagent/Resource | Specifications | Function/Application | Example Sources |
|---|---|---|---|
| TRV Vectors | pTRV1, pTRV2 with multiple cloning sites | Viral replication and insert delivery | [54] [5] |
| Agrobacterium Strain | GV3101 with pSoup helper plasmid | Delivery of T-DNA to plant cells | [54] [27] |
| Selection Antibiotics | Kanamycin, Rifampicin | Selection of transformed Agrobacterium | [54] [27] |
| Induction Media | YEB with MES, Acetosyringone | Induction of vir genes for T-DNA transfer | [54] [27] |
| DNA Methylation Analysis Kits | Bisulfite conversion kits | Detection of DNA methylation patterns | [1] |
| siRNA Detection Reagents | Northern blot or sequencing kits | Verification of siRNA production | [1] [53] |
| Plant Growth Regulators | Specific to plant species | Enhanced transformation efficiency | Species-dependent |
Technical Considerations and Limitations
Despite its promise, VIGS-induced epigenetic silencing presents several technical challenges that researchers must consider:
Temperature Sensitivity: Standard TRV-based VIGS systems function optimally between 19-25°C, with efficiency dramatically reduced at higher temperatures [55]. Recent development of California TRV isolates with enhanced thermal tolerance (maintaining 78% efficiency at 30°C) partially addresses this limitation but may be associated with increased growth stunting phenotypes [55].
Species-Specific Optimization: The efficiency of VIGS-induced epigenetics varies significantly among plant species, necessitating optimization of multiple parameters including:
- Infiltration method
- Plant developmental stage
- Agrobacterium density
- Fragment length and specificity [54] [27]
Stability and Penetrance: The establishment of heritable epigenetic marks is probabilistic rather than deterministic, resulting in variable silencing penetrance across treated plants and potential instability in subsequent generations. Factors influencing stability include:
- Target sequence composition (C-residue density)
- Efficiency of RNA-dependent and independent maintenance mechanisms
- Developmental resetting of epigenetic marks [1]
The integration of VIGS with emerging technologies promises to enhance its application in epigenetic research and crop improvement:
Combination with Genome Editing: The fusion of VIGS with CRISPR-dCas9 systems enables targeted epigenetic modifications without altering DNA sequences, offering precise control over gene expression [1] [56].
High-Throughput Functional Genomics: VIGS-based epigenetic screening platforms facilitate large-scale identification of genes regulating agronomic traits, accelerating functional genomics in species resistant to stable transformation [3] [27].
Field Application Development: The creation of temperature-tolerant VIGS vectors enables field studies of gene function under natural conditions, particularly valuable for ecological and agricultural research [55].
In conclusion, VIGS technology has transcended its original application as a transient gene silencing tool to become a powerful platform for inducing stable, transgenerational epigenetic modifications. By leveraging the plant's native RdDM machinery, researchers can now create novel epialleles with potential applications in crop improvement, synthetic biology, and fundamental research on epigenetic inheritance. While technical challenges remain, ongoing optimization of vector systems, delivery methods, and species-specific protocols continues to expand the utility of this technology across diverse plant systems.
Maximizing Silencing Efficiency: A Guide to Troubleshooting and Protocol Optimization
Virus-Induced Gene Silencing (VIGS) has emerged as a cornerstone technology for reverse genetics, enabling rapid functional analysis of plant genes by harnessing the plant's innate RNA interference (RNAi) machinery. This process involves using recombinant viral vectors to deliver host-derived gene fragments, triggering sequence-specific mRNA degradation and resulting in targeted gene knockdown [3]. While VIGS is well-established in model plants like Nicotiana benthamiana and Arabidopsis thaliana, its application in recalcitrant species—particularly perennial woody plants—presents significant challenges. These species often exhibit physical barriers (e.g., lignified tissues), sophisticated antiviral defense mechanisms, and inefficient viral movement, which collectively contribute to host resistance and limit VIGS efficacy [3] [27].
The imperative to overcome this resistance is driven by the growing need to functionally characterize genes in agronomically valuable woody crops, many of which are poorly amenable to stable transformation and traditional genomic editing techniques [3] [27]. This technical guide synthesizes recent advances in VIGS vector engineering, delivery optimization, and experimental design, providing a strategic framework for implementing robust gene silencing systems in recalcitrant and woody plant species within the broader context of VIGS RNA-mediated reverse genetics research.
Core Principles and Challenges of VIGS in Recalcitrant Species
The biological foundation of VIGS lies in the plant's post-transcriptional gene silencing (PTGS) pathway, a conserved antiviral defense mechanism. Upon introduction, recombinant viral vectors containing sequences homologous to a plant gene of interest (GOI) replicate and produce double-stranded RNA (dsRNA). This dsRNA is recognized and cleaved by host Dicer-like (DCL) enzymes into 21- to 24-nucleotide small interfering RNAs (siRNAs). These siRNAs are then incorporated into an RNA-induced silencing complex (RISC), which guides the sequence-specific degradation of complementary endogenous mRNA transcripts, leading to a loss-of-function phenotype [3] [13].
In recalcitrant and woody species, several interconnected challenges can impede this process:
- Physical and Developmental Barriers: Dense, lignified tissues in organs like mature stems and fruit capsules can physically obstruct viral entry and systemic movement. Furthermore, the slow growth and long life cycles of woody perennials extend experimental timelines and can dilute silencing signals over time [27].
- Enhanced Antiviral Defenses: Some species possess highly efficient RNAi machinery or dominant resistance genes that rapidly contain or eliminate viral infections, thereby curtailing the spread and persistence of the VIGS vector [3] [13].
- Inefficient Meristem Invasion: A key hurdle for achieving heritable edits via viral delivery is the plant's ability to restrict viruses from invading meristematic and germline tissues, which is a common feature in many species [57].
The following diagram illustrates the core mechanism of VIGS and the major points of host resistance in recalcitrant plants.
Strategic Optimization of VIGS Components
Advanced Vector Engineering and Selection
Choosing and engineering the appropriate viral vector is the most critical determinant of VIGS success in recalcitrant hosts.
- Vector Selection Based on Host Range: The Tobacco Rattle Virus (TRV) is one of the most versatile and widely used VIGS vectors, especially within the Solanaceae family, due to its broad host range, efficient systemic movement, and ability to target meristematic tissues [3]. For species outside the TRV host range, exploring vectors based on viruses that naturally infect related species is essential. The 30K family of movement proteins (MPs), which includes over 500 viral species capable of infecting all agronomically important plants, provides a rich resource for vector development [31].
- Movement Protein (MP) Engineering: A novel approach involves modifying the coding sequence of viral movement proteins themselves to carry small inserts (as short as 18-54 bp) of the host target gene. This strategy, successfully demonstrated with Alfalfa mosaic virus (AMV), Cucumber mosaic virus (CMV), and Tobacco mosaic virus (TMV), can bypass compatibility issues and is applicable to the entire 30K MP family [31].
- Employing Ultra-Compact Systems: For enabling transgene-free germline editing, recent research has engineered TRV to carry ultra-compact RNA-guided editors like the TnpB enzyme (~400 amino acids). This system overcame traditional cargo capacity limits of viral vectors and achieved heritable edits in Arabidopsis, representing a significant breakthrough for functional genomics [57].
- Utilization of Viral Suppressors of RNA Silencing (VSRs): Co-expressing well-characterized VSRs like P19 or HC-Pro can transiently suppress the host's antiviral RNAi machinery, thereby enhancing viral accumulation and spreading the silencing signal. However, this must be carefully calibrated to avoid severe plant toxicity [3].
Table 1: Key Viral Vectors for Recalcitrant and Woody Species
| Vector System | Virus Type | Key Features | Demonstrated Hosts/Applications |
|---|---|---|---|
| Tobacco Rattle Virus (TRV) | RNA Virus (Bipartite) | Broad host range; efficient systemic movement; targets meristem tissues [3]. | Nicotiana benthamiana, Capsicum annuum (Pepper), Camellia drupifera [3] [27]. |
| 30K Family MP Vectors | Various (AMV, CMV, TMV) | MP modified to carry small inserts (18-54 bp); wide host range across agronomic species [31]. | N. tabacum, N. benthamiana; potentially all hosts of the 30K MP family [31]. |
| TRV-TnpB Editor | RNA Virus (Bipartite) | Carries compact TnpB nuclease for germline editing; enables transgene-free, heritable mutations [57]. | Arabidopsis thaliana (Proof-of-concept) [57]. |
Optimized Delivery Methodologies
The method of viral vector delivery must be tailored to the specific morphology and tissue architecture of the woody plant.
- Agroinfiltration Parameters: Standard leaf infiltration is often ineffective. For fruits and capsules, methods like pericarp cutting immersion and direct pericarp injection have proven highly effective. In Camellia drupifera, pericarp cutting immersion achieved an infiltration efficiency of ~94% [27]. The optical density (OD600) of the agrobacterium inoculum (typically 0.9-1.0), the use of acetosyringone to induce virulence genes, and the developmental stage of the tissue are all critical parameters that require optimization [27].
- Developmental Stage Timing: Silencing efficiency is profoundly influenced by the developmental stage of the target organ. Research in C. drupifera capsules showed that the optimal VIGS effect for different genes was achieved at specific stages: early development (~70% efficiency for CdCRY1) and mid-stage (~91% efficiency for CdLAC15) [27]. Targeting tissues during active growth phases before extensive lignification occurs is a key strategy.
Table 2: Comparison of Delivery Methods for Recalcitrant Tissues
| Delivery Method | Procedure | Advantages | Limitations | Reported Efficiency |
|---|---|---|---|---|
| Pericarp Cutting Immersion | Fruit capsule is cut and immersed in agrobacterium solution [27]. | High efficiency; allows direct access to susceptible cells. | Limited to specific organ types; causes physical damage. | ~94% infiltration efficiency in C. drupifera [27]. |
| Direct Pericarp Injection | Agrobacterium suspension is injected directly into the fruit pericarp [27]. | Targets specific tissue layers; good for deeper tissues. | Technically demanding; potential for clogging. | Quantified in specific varieties of C. drupifera [27]. |
| Peduncle Injection | Inoculum is delivered via the fruit stalk (peduncle) [27]. | Utilizes vascular system for distribution. | May be less efficient than direct pericarp methods. | Evaluated in C. drupifera [27]. |
| Agroflood | Soil-drench inoculation of whole seedlings [57]. | Less labor-intensive; suitable for high-throughput screening. | May be less efficient in mature plants with robust root systems. | Used for TRV-TnpB delivery in Arabidopsis [57]. |
Insert and Environmental Optimization
- Insert Design for Specificity and Efficiency: The standard insert size for VIGS is 200-500 bp. However, using shorter, highly specific sequences (e.g., 21-45 bp) can reduce off-target effects while still inducing effective silencing. A clear correlation exists between insert size and silencing efficacy, with fragments of 45 nucleotides or larger achieving 75-90% efficiency [31] [58]. Tools like the SGN VIGS Tool should be used to screen for sequence specificity and minimize homology with non-target genes [27].
- Control of Environmental Conditions: Cultivation conditions post-inoculation significantly impact silencing robustness. Temperature is a critical factor; for instance, studies with the TRV-TnpB system in Arabidopsis showed that heat treatment could increase editing efficiency by over 6-fold [57]. Maintaining optimal humidity and photoperiod is also essential to support plant health and viral spread without triggering hypersensitive defense responses [3].
Integrated Experimental Workflow and Toolkit
The following diagram and table provide a consolidated overview of the experimental journey and essential reagents for establishing VIGS in a recalcitrant species.
Table 3: The Scientist's Toolkit: Essential Reagents for VIGS in Recalcitrant Plants
| Reagent / Material | Function / Purpose | Specifications & Notes |
|---|---|---|
| pTRV1 & pTRV2 Vectors | Bipartite TRV system; TRV1 encodes replication proteins, TRV2 carries the gene insert [3]. | Most widely used system; available from multiple academic repositories. |
| Modified MP Vectors (e.g., pAMV-MP) | Viral vectors engineered to carry small inserts within the Movement Protein gene [31]. | Ideal for fine-tuning silencing levels with inserts <100 bp. |
| Agrobacterium tumefaciens | Bacterial vehicle for delivering viral vectors into plant cells. | Strains GV3101 and LBA4404 are commonly used. |
| Acetosyringone | Phenolic compound that induces the Agrobacterium virulence genes. | Critical for enhancing transformation efficiency; used in induction medium [27]. |
| Selection Antibiotics | Maintains plasmid stability in bacterial and plant cultures. | e.g., Kanamycin (25-50 μg/mL), Rifampicin (50 μg/mL) [27]. |
| Infiltration Buffers | Medium for resuspending agrobacteria for inoculation. | Typically contains MgCl₂, MES, and acetosyringone. |
| Visual Marker Genes | Allows for rapid assessment of infiltration success and silencing spread. | e.g., Green Fluorescent Protein (GFP) cloned into TRV2 vector [27]. |
| Endogenous Marker Genes | Positive control for silencing efficiency based on visible phenotype. | e.g., Phytoene Desaturase (PDS), causing photobleaching [3] [31]. |
The successful application of VIGS in recalcitrant and woody plant species is no longer an insurmountable challenge but a manageable process of strategic optimization. The convergence of advanced vector engineering—including MP-modified viruses and compact editing systems—with tailored delivery methods and precise environmental control provides a powerful toolkit for researchers. By systematically addressing the unique physiological and biochemical barriers presented by these hosts, as demonstrated in successful case studies like Camellia drupifera, scientists can leverage VIGS to accelerate functional genomics and trait validation in a wide range of previously intractable species. This progress is pivotal for advancing reverse genetics research and accelerating the breeding of next-generation, climate-resilient woody crops.
Virus-Induced Gene Silencing (VIGS) has established itself as a powerful reverse genetics tool, enabling researchers to rapidly characterize gene function by leveraging the plant's innate RNA interference (RNAi) machinery [3] [1]. The efficacy of this technology hinges on the strategic design of the insert—the fragment of the target plant gene incorporated into the viral vector. A well-designed insert ensures efficient, specific, and stable knockdown of the gene of interest, whereas poor design can lead to incomplete silencing, unintended effects, or rapid loss of the insert from the viral genome [45]. This guide details the core principles of VIGS insert design—optimizing length, ensuring specificity, and mitigating off-target effects—providing a foundational framework for robust functional genomics research.
Core Design Parameters for VIGS Inserts
The strategic design of the DNA fragment inserted into the VIGS vector is paramount to the experiment's success. The following parameters must be carefully balanced to achieve effective and specific gene silencing.
Insert Length
The length of the inserted gene fragment is a critical determinant of silencing efficiency and stability.
- Optimal Range: Inserts typically between 200 and 500 base pairs (bp) are recommended for effective silencing [3] [27]. This size is sufficient to trigger a strong RNAi response while being manageable for the viral vector to replicate and transport systemically.
- Practical Application: In a study developing a VIGS system for the recalcitrant woody plant Camellia drupifera, researchers successfully used inserts of 200-300 bp to silence genes involved in pericarp pigmentation, achieving high efficiency [27].
- Stability Considerations: While shorter inserts may be replicated more efficiently, they might generate an insufficient number of siRNAs. Excessively long inserts can impose a replicative burden on the virus, leading to genomic instability and the deletion of the foreign sequence over time [45].
Table 1: VIGS Insert Length Guidelines
| Insert Length | Advantages | Disadvantages | Recommended Use |
|---|---|---|---|
| 200 - 500 bp | Optimal balance of silencing efficiency and viral stability [3] [27] | May still be unstable in some vector systems | Standard, robust experimental design |
| < 200 bp | Potentially less burdensome on the virus | May produce insufficient siRNAs, leading to weak silencing | Less common; requires empirical validation |
| > 500 bp | May generate a diverse siRNA pool | High probability of deletion and vector instability [45] | Generally not recommended |
Sequence Specificity and Selection
Ensuring the insert sequence uniquely corresponds to the intended target gene is essential to avoid unintentionally silencing related genes.
- Homology Analysis: Before constructing the vector, the candidate insert sequence must be analyzed for homology against the host plant's genome or transcriptome databases. Tools like the SGN VIGS Tool are available for this purpose [27].
- Specificity Threshold: A common practice is to select a fragment with less than 40% sequence similarity to other genes in the genome, thereby minimizing the risk of cross-silencing non-target genes [27].
- Target Region: Inserts are typically designed from the coding sequence (CDS) of the target gene [1]. For the specific aim of inducing transcriptional gene silencing (TGS) via promoter methylation, the insert must correspond to the promoter region of the target gene instead [1].
Avoiding Off-Target Effects
Off-target effects occur when siRNAs derived from the VIGS vector bind to and silence mRNAs of other genes with partial sequence complementarity.
- Bioinformatic Screening: The most crucial step is rigorous in silico analysis. Potential siRNA sequences generated from the candidate insert should be checked for complementarity to other genes in the host genome, focusing on seed regions. This process is facilitated by various target prediction software tools [59].
- Validation with Controls: The phenotypic consequences of VIGS should be validated using multiple, independent inserts targeting different regions of the same gene. This ensures that the observed phenotype is due to the knockdown of the intended gene and not an off-target effect [59].
Advanced Consideration: Ensuring Insert Stability
A cutting-edge challenge in VIGS, particularly for long-term applications in perennial plants, is the tendency of viral vectors to expel foreign inserts through recombination during replication [45].
Recent research indicates that insert stability is not merely a function of sequence or length but is profoundly influenced by the thermodynamic properties of the inserted RNA secondary structure. A 2025 study on the Citrus yellow vein-associated virus (CY1) vector demonstrated that foreign hairpins with thermodynamic properties (e.g., positional entropy and free energy, ΔG) differing from those of natural viral hairpin structures were rapidly deleted [45]. In contrast, inserts designed to mimic the thermodynamic profile of native viral hairpins were stably maintained for over 30 months in citrus plants [45].
Practical Implication: For vectors with known RNA secondary structures, designing inserts that conform to the virus's innate structural and thermodynamic landscape can dramatically improve stability. This involves computational prediction of RNA folding and thermodynamic parameters to guide insert design [45].
Experimental Protocol: A Workflow for VIGS Insert Design and Validation
The following protocol outlines the key steps for designing, constructing, and validating a VIGS construct.
Step-by-Step Procedures
- Target Identification and Sequence Retrieval: Obtain the full coding sequence (CDS) of your target gene from a genomic or transcriptomic database.
- In Silico Specificity Screening:
- Fragment Selection and Design:
- Design PCR primers with appropriate restriction enzyme sites for cloning into your VIGS vector (e.g., pTRV2).
- For advanced stability, use RNA folding prediction software (e.g., RNAfold) to analyze the thermodynamic properties (ΔG, positional entropy) of the proposed insert. If possible, model it after stable hairpins in the viral genome itself [45].
- Cloning and Transformation:
- Amplify the fragment via PCR from cDNA using high-fidelity DNA polymerase.
- Ligate the purified PCR product into the VIGS vector using a method like Nimble Cloning.
- Transform the recombinant plasmid into E. coli DH5α competent cells for propagation.
- Sequence several positive colonies to confirm the insert is correct and in the right orientation [27].
- Plant Inoculation:
- Transform the validated plasmid into Agrobacterium tumefaciens (e.g., strain GV3101).
- Prepare agrobacteria cultures containing the VIGS vector (e.g., TRV2 with insert) and the helper vector (e.g., TRV1). Induce with acetosyringone.
- For petunia, inoculating mechanically wounded shoot apical meristems is highly effective [60]. For cotton and other species, cotyledon agroinfiltration is standard [28] [27]. Methods like pericarp cutting immersion are optimized for recalcitrant tissues [27].
- Molecular Validation:
- RNA Extraction: Isolate total RNA from silenced tissues using a commercial kit (e.g., Spectrum Total RNA Extraction Kit) [28].
- Reverse Transcription: Synthesize first-strand cDNA.
- RT-qPCR:
- Use validated reference genes that are stable under VIGS conditions and experimental stresses. A 2025 study in cotton under aphid herbivory stress found GhACT7 and GhPP2A1 to be superior, whereas commonly used genes like GhUBQ7 were unstable and led to inaccurate results [28].
- Design qPCR primers outside the region targeted by the VIGS insert to avoid amplification of the viral RNA.
- Calculate relative expression using the ΔΔCt method normalized to the stable reference genes. Successful knockdown is typically indicated by a significant reduction (e.g., >50%) in target mRNA levels [28].
The Scientist's Toolkit: Essential Reagents for VIGS
Table 2: Key Research Reagents and Solutions for VIGS Experiments
| Reagent / Material | Function / Description | Example(s) |
|---|---|---|
| Viral Vectors | Engineered viruses to deliver the insert; bipartite for TRV. | pTRV1 (Helper), pTRV2 (with MCS for insert) [3] [27] |
| Agrobacterium Strain | Delivery vehicle for the viral DNA construct. | A. tumefaciens GV3101 [28] [27] |
| Induction Buffer | Activates Agrobacterium for plant infection. | 10 mM MES, 10 mM MgCl₂, 200 µM Acetosyringone [28] |
| Stable Reference Genes | Critical for accurate RT-qPCR normalization under VIGS conditions. | GhACT7, GhPP2A1 (in cotton) [28] |
| Visual Marker Genes | Positive controls to visually monitor silencing efficiency. | PDS (photo-bleaching), CHS (loss of pigment) [3] [60] |
| Control VIGS Vector | Essential control for viral symptoms and non-specific effects. | pTRV2 with non-plant insert (e.g., GFP) [60] |
Molecular Mechanisms of VIGS and Insert Function
The designed insert is central to triggering the plant's RNAi machinery. The following diagram illustrates the key molecular steps from agroinfiltration to gene silencing.
The precision of VIGS as a functional genomics tool is fundamentally dictated by the principles of insert design. Adherence to the guidelines for length (200-500 bp), rigorous bioinformatic screening for specificity, and careful consideration of advanced factors like thermodynamic stability for insert retention, collectively form the foundation of a successful experiment. By integrating these design principles with robust experimental protocols and appropriate controls—including the use of stable reference genes for validation—researchers can minimize artifacts and confidently attribute phenotypes to the knockdown of their gene of interest. As VIGS technology continues to evolve, these refined design strategies will be instrumental in unraveling complex gene functions and accelerating reverse genetics research.
Agroinfiltration is a cornerstone technique in modern plant biotechnology, enabling the transient delivery of genetic material into plant tissues for a wide range of applications, from recombinant protein production to functional genomics. Within the broader context of Virus-Induced Gene Silencing (VIGS) research—an RNA-mediated reverse genetics technology—the precision of agroinfiltration is paramount. VIGS leverages the plant's innate post-transcriptional gene silencing (PTGS) machinery, activated by recombinant viral vectors delivered via Agrobacterium tumefaciens, to knock down target gene expression and characterize gene function [1] [3]. The efficacy of this process is highly dependent on the physiological interactions between the bacterial inoculum and the host plant tissue. Key technical parameters, specifically the optical density at 600 nm (OD600) of the Agrobacterium culture, the concentration of the phenolic elicitor acetosyringone, and the developmental stage of the plant material, are critical determinants of successful T-DNA transfer and subsequent transgene expression or silencing. This guide synthesizes current research to provide a detailed, evidence-based framework for optimizing these parameters to achieve robust, reproducible results in VIGS and other agroinfiltration-based assays.
The Molecular Interplay of Agroinfiltration and VIGS
Optimizing agroinfiltration for VIGS requires an understanding of the synergistic molecular pathways activated in both the bacterial vector and the host plant. The diagram below illustrates the key processes from bacterial induction to the establishment of gene silencing.
The diagram above outlines the core workflow. Acetosyringone in the infiltration medium activates the bacterial vir genes, initiating T-DNA transfer [61] [62]. The developmental stage of the plant tissue influences the efficiency of bacterial infection and the systemic spread of the silencing signal [27] [63]. Once delivered, the viral vector replicates, leading to double-stranded RNA (dsRNA) formation. This dsRNA is cleaved by Dicer-like (DCL) enzymes into small interfering RNAs (siRNAs) that guide the RNA-induced silencing complex (RISC) to degrade complementary mRNA sequences (PTGS), resulting in transient gene knockdown [1] [3]. In some cases, siRNAs can also lead to transcriptional gene silencing (TGS) via RNA-directed DNA methylation (RdDM) [1].
Quantitative Optimization Parameters
Optimizing agroinfiltration is a multivariable process. The table below consolidates empirically determined optimal ranges for key parameters across various plant species.
Table 1: Optimal agroinfiltration parameters for different plant systems in VIGS and transient expression.
| Plant System | Optimal OD₆₀₀ | Optimal Acetosyringone Concentration | Optimal Plant Developmental Stage / Tissue | Key Supporting Findings |
|---|---|---|---|---|
| Nicotiana benthamiana (General Transfection) | 0.1 - 1.5 [61] [62] | 150 – 500 µM [61] [62] | 4-6 week old plants; expanded leaves [62] | 500 µM acetosyringone significantly increased GUS reporter expression [61]. |
| Soybean (Glycine max) (TRV-VIGS) | ~0.5 [7] | 200 µM [7] | Cotyledon node of half-seed explants [7] | Use of half-seed explants achieved up to 95% infection efficiency [7]. |
| Tea Plant (Camellia sinensis) (TRV-VIGS) | 0.9 - 1.0 [63] | 200 µM [63] | Young cuttings (8-10 cm) with new buds [63] | Highest CsPDS silencing efficiency (63.3%) achieved in young, actively growing cuttings. |
| Camellia drupifera (TRV-VIGS) | 0.9 - 1.0 [27] | 100 µM [27] | Early to mid-stage capsules (279 days post-pollination) [27] | Silencing efficiency was ~70-91%, dependent on capsule developmental stage and target gene. |
| Mulberry (Morus L.) (Transient Transf.) | 0.5 [64] | 150 µM [64] | Leaves from 4-week-old tissue culture seedlings [64] | Combined with sonication/vacuum, this yielded high GFP expression in 'Taiguo' genotype. |
Interpreting the Data and Practical Implications
- OD600: The data reveals a common optimal OD600 range between 0.5 and 1.0 for many systems [61] [7] [64]. Lower densities may result in insufficient T-DNA delivery, while higher densities can trigger a hypersensitive defense response in the plant, leading to tissue necrosis and reduced gene expression [61].
- Acetosyringone: This phenolic compound is essential for inducing the Agrobacterium vir genes. While a concentration of 150-200 µM is widely used and effective [64] [62] [63], studies in N. benthamiana have shown that increasing the concentration to 500 µM can provide a significant boost to transient expression levels [61].
- Plant Developmental Stage: The most consistent finding is that young, metabolically active tissues are most amenable to agroinfiltration and VIGS. In tea plants, silencing was only observed in newly emerged buds, not in mature leaves [63]. Similarly, in soybean, using the meristematic cells of the cotyledon node was critical for systemic TRV spread [7]. For specialized tissues like C. drupifera capsules, the developmental window is narrow and must be empirically determined [27].
Experimental Protocol for an Optimized VIGS Assay
Below is a detailed methodology for conducting a TRV-based VIGS experiment in a typical plant system, incorporating the optimized parameters discussed.
Agrobacterium Culture and Inoculum Preparation
- Transform and Select Agrobacterium: Introduce the binary VIGS vectors (e.g., pTRV1 and pTRV2 with target gene insert) into a suitable Agrobacterium strain (e.g., GV3101 or LBA4404) [7] [64].
- Starter Culture: Inoculate a single bacterial colony into 5-10 mL of LB medium with appropriate antibiotics (e.g., kanamycin, rifampicin) and incubate at 28°C with shaking for 24-48 hours [27].
- Induction Culture: Dilute the starter culture 1:50 into a fresh induction medium (e.g., YEB or LB with 10 mM MES, pH 5.6, and 200 µM acetosyringone). Grow the culture at 28°C with shaking until it reaches the target OD600 of 0.5 to 1.0 [27] [63].
- Harvest and Resuspend: Pellet the bacterial cells by centrifugation (e.g., 5000 rpm for 15 minutes). Gently resuspend the pellet in an infiltration medium (e.g., 10 mM MgCl₂, 10 mM MES, pH 5.6, and 150-200 µM acetosyringone) to the final working OD600 [7] [62].
- Induce Virulence: Incubate the resuspended agrobacteria at room temperature for 3-6 hours without shaking to fully activate the vir genes [62].
Plant Material Preparation and Infiltration
- Select Plant Material: Use plants at the optimal developmental stage. For many species, this is the young seedling stage or plants with newly expanded, tender leaves [7] [63].
- Mix Agrobacterial Suspensions: Combine the resuspended pTRV1 and pTRV2 cultures in a 1:1 ratio immediately before infiltration [62].
- Infiltrate Plant Tissue:
- For leaves of model plants like N. benthamiana: Use a needleless syringe to press against the abaxial (lower) side of the leaf, gently infiltrating the suspension [62].
- For recalcitrant tissues or high-throughput: Employ optimized methods such as vacuum infiltration (e.g., 0.8 kPa for 5 minutes for tea plants) [63] or pericarp cutting immersion (for woody capsules) [27].
The Scientist's Toolkit: Essential Research Reagents
A successful agroinfiltration experiment relies on a suite of carefully selected reagents. The following table details the core components and their specific functions.
Table 2: Key reagents for agroinfiltration and VIGS experiments.
| Reagent / Material | Function / Role | Typical Working Concentration |
|---|---|---|
| Acetosyringone | Phenolic elicitor that activates Agrobacterium vir genes, essential for T-DNA transfer. | 150 - 500 µM [61] [62] |
| MES Buffer | Maintains optimal pH (5.6) of the infiltration medium to stabilize Agrobacterium and support vir gene induction. | 10 mM [64] [62] |
| MgCl₂ | Provides essential divalent cations, maintaining osmotic balance and bacterial membrane integrity. | 10 mM [62] |
| Silwet L-77 Surfactant | Reduces surface tension, allowing the infiltration medium to spread through intercellular spaces, boosting efficiency. | 0.002% - 0.02% [61] [64] |
| Agrobacterium Strain GV3101 | A disarmed, helper plasmid-containing strain widely used for its high transformation efficiency in many dicots. | N/A [7] [37] |
| TRV-based VIGS Vectors | Deconstructed viral vector system (pTRV1, pTRV2) known for broad host range and efficient systemic silencing with mild symptoms. | N/A [7] [3] [27] |
The meticulous optimization of OD600, acetosyringone concentration, and plant developmental stage is not merely a procedural step but a fundamental requirement for harnessing the full potential of agroinfiltration in VIGS research. By aligning bacterial physiology with plant cell receptivity, researchers can achieve high-efficiency gene silencing. The parameters and protocols detailed in this guide provide a robust foundation for advancing reverse genetics studies, ultimately accelerating the functional annotation of plant genomes and the development of crops with improved agronomic traits. As VIGS technology continues to evolve, integrating these optimized physical and chemical parameters with novel vector designs and delivery methods will further solidify its role as an indispensable tool in plant functional genomics and biotechnology.
Virus-Induced Gene Silencing (VIGS) is an RNA-mediated reverse genetics technique that has evolved into an indispensable approach for analyzing gene function in plants by downregulating endogenous genes through the plant's post-transcriptional gene silencing (PTGS) machinery [1]. The effectiveness of VIGS is not solely dependent on the molecular design of viral vectors but is profoundly influenced by the environmental conditions in which the experimental plants are maintained [3] [65]. Controlling environmental factors such as temperature, humidity, and photoperiod is therefore not merely a matter of plant husbandry but a critical determinant in the success and reproducibility of VIGS experiments. This technical guide synthesizes current knowledge and protocols to empower researchers to master these environmental variables, thereby ensuring robust and reliable gene silencing outcomes.
The Critical Role of Environmental Factors in VIGS Efficiency
The VIGS process leverages the plant's innate antiviral defense system. When a recombinant virus carrying a fragment of a host gene is introduced, the plant's RNA silencing machinery is activated, leading to sequence-specific degradation of homologous endogenous mRNA transcripts [1] [66]. This cellular machinery, involving Dicer-like enzymes and RNA-induced silencing complexes (RISC), is highly sensitive to the plant's physiological status, which is in turn modulated by its environment [3].
Suboptimal environmental conditions can negatively impact VIGS efficiency through multiple mechanisms:
- Weakened Plant Vigor: Stressful conditions can reduce the overall health and metabolic activity of the plant, impeding the systemic spread of the viral vector and the silencing signal [65].
- Altered Viral Replication and Movement: Temperature and light directly affect viral replication rates and the function of viral movement proteins, influencing the speed and extent of infection [67].
- Impaired Silencing Machinery: The enzymatic activities central to PTGS, including Dicers and Argonautes, are temperature-sensitive. Furthermore, the intercellular and long-distance movement of siRNAs exhibits species-specific variation that can be influenced by environmental factors [3].
Mastery of the environment allows researchers to standardize these variables, turning them from sources of experimental noise into levers for optimizing silencing efficacy.
Quantitative Guidelines for Key Environmental Parameters
A synthesis of research across multiple plant species provides quantitative guidance for the optimal control of temperature, humidity, and photoperiod. The following table summarizes key parameters for different experimental contexts.
Table 1: Optimal Environmental Parameters for VIGS Across Plant Species and Conditions
| Plant Species | Temperature | Photoperiod | Humidity | Key Effect on VIGS | Primary Citation |
|---|---|---|---|---|---|
| General Greenhouse Setting | ~22 °C (average) | 16-h light / 8-h dark (Long-day) | ~45% (relative) | Promotes vigorous plant growth and efficient systemic silencing. | [65] |
| Arabidopsis thaliana (Optimal) | Not Specified | 16-h light / 8-h dark (Long-day) | Not Specified | 90-100% of plants showed silencing; crucial for high efficiency. | [68] |
| Arabidopsis thaliana (Suboptimal) | Not Specified | 8-h light / 16-h dark (Short-day) | Not Specified | Only 10% of plants showed silencing. | [68] |
| Nicotiana benthamiana (Post-Inoculation) | 21±2 °C | Not Specified | Not Specified | Maintains vigorous growth, which is critical for efficient VIGS. | [66] |
Experimental Protocols for Environmental Optimization
Protocol: Establishing Baseline Conditions forArabidopsis thaliana
This protocol, adapted from Burch-Smith et al. (2006), is fundamental for achieving high VIGS efficiency in the model organism Arabidopsis thaliana ecotype Columbia-0 (Col-0) using TRV vectors [68].
Plant Growth and Selection:
- Sow seeds in a suitable soil-less potting mixture.
- Grow seedlings under long-day conditions (16-h light / 8-h dark photoperiod).
- Select plants at the two- to three-leaf stage for agroinfiltration. Using older plants (e.g., four- to five-leaf stage) can reduce silencing efficiency by up to 50%.
Agroinfiltration:
- Resuspend Agrobacterium tumefaciens cultures carrying TRV1 and TRV2 vectors to an optical density at 600 nm (OD~600~) of 1.5 in inoculation buffer (e.g., 10 mM MES, pH 5.5; 200 μM acetosyringone).
- Mix the cultures in a 1:1 ratio and incubate for several hours.
- Infiltrate the mixture into the abaxial side of leaves using a needleless syringe.
Post-Inoculation Care:
- Return infiltrated plants to the long-day growth chamber.
- Maintain consistent temperature and provide adequate nutrition and water to support vigorous growth.
- Silencing phenotypes, such as photobleaching from PDS silencing, typically become visible within two to three weeks post-inoculation.
Protocol: Advanced Seed-Vacuum Infiltration for Recalcitrant Species
For species that are difficult to transform, such as sunflower, a seed-vacuum method has been developed to achieve high infection and silencing rates [65].
Plant Material Preparation:
- Partially remove the seed coat ("peeling") to facilitate infiltration without damaging the embryo. No surface sterilization or in vitro recovery is needed.
Agrobacterium Preparation:
- Transform TRV constructs (pTRV1 and pTRV2 with gene insert) into Agrobacterium (e.g., strain GV3101).
- Initiate cultures from single colonies on selective LB-agar plates.
- Inoculate liquid cultures and grow to desired density. Harvest cells by centrifugation and resuspend in infiltration medium (e.g., 10 mM MgCl~2~, 10 mM MES, 150 μM acetosyringone) to a final OD~600~ of ~1.5-2.0.
Vacuum Infiltration and Co-cultivation:
- Submerge the peeled seeds in the Agrobacterium suspension in a vacuum desiccator.
- Apply a vacuum (e.g., 0.8-0.9 Bar) for 5-10 minutes, then release rapidly to allow the suspension to penetrate the seeds.
- Pour the suspension with seeds into a sterile container for co-cultivation. Place the container on a shaker (50-100 rpm) for 6 hours at room temperature in the dark.
Post-Infection Plant Cultivation:
- Sow the treated seeds directly into a peat-perlite (3:1) mixture in pots.
- Transfer pots to a greenhouse maintained at an average of 22°C, 45% relative humidity, and an 18-h light / 6-h dark photoperiod.
- Silencing symptoms in newly developed leaves can be expected 2-3 weeks post-treatment.
Visualizing the Environmental Impact on VIGS Efficiency
The diagram below illustrates the logical relationship between controlled environmental inputs, the resulting plant physiological state, and the ultimate effect on key VIGS processes.
The Scientist's Toolkit: Essential Reagents and Materials
The following table details key reagents and materials required for implementing the environmental-controlled VIGS protocols described in this guide.
Table 2: Essential Research Reagent Solutions for VIGS Experiments
| Reagent/Material | Function/Description | Example Use Case |
|---|---|---|
| TRV Vectors (pTRV1, pTRV2) | Bipartite viral vector system; TRV1 encodes replication proteins, TRV2 carries the target gene insert for silencing. | The most widely used VIGS system for Solanaceous species and Arabidopsis [68] [3]. |
| Agrobacterium tumefaciens | Bacterial vehicle for delivering TRV vectors into plant cells via T-DNA transfer. Strain GV3101 is commonly used. | Used in both standard agroinfiltration and seed-vacuum protocols [68] [65]. |
| Infiltration Buffer | Buffer for suspending Agrobacterium; typically contains MgCl₂, MES, and acetosyringone. | Acetosyringone induces vir gene expression, enhancing T-DNA transfer during agroinfiltration [66] [65]. |
| Selective Antibiotics | Kanamycin, rifampicin, gentamicin; for selecting and maintaining bacterial strains with TRV plasmids. | Ensuring plasmid retention in Agrobacterium cultures during preparation [65]. |
| Phytoene Desaturase (PDS) Clone | A positive control marker gene; silencing PDS causes photobleaching (white patches), visually confirming VIGS efficiency. | Standard marker for optimizing environmental parameters and protocols [68] [65]. |
The precise control of temperature, humidity, and photoperiod is a fundamental, yet sometimes overlooked, component of robust VIGS experimental design. By adhering to the quantitative guidelines, detailed protocols, and reagent specifications outlined in this technical guide, researchers can significantly enhance the efficiency, penetration, and reproducibility of virus-induced gene silencing. As VIGS continues to be a cornerstone technology in plant functional genomics and crop improvement [1] [3], mastering the plant's environment ensures that the observed phenotypes are a true reflection of gene function rather than an artifact of suboptimal growth conditions.
Employing Viral Suppressors of RNA Silencing (VSRs) to Enhance Silencing Efficacy
RNA silencing is a conserved, sequence-specific gene regulation mechanism that serves as a crucial antiviral defense system in plants and other eukaryotic organisms. This system processes double-stranded RNA (dsRNA) into 21-24 nucleotide small interfering RNAs (siRNAs) that guide the RNA-induced silencing complex (RISC) to cleave complementary viral RNA sequences [69]. To counteract this host defense, most plant viruses have evolved viral suppressors of RNA silencing (VSRs) - diverse proteins that inhibit various stages of the silencing pathway [70]. The strategic application of these VSRs has emerged as a powerful approach to enhance the efficacy of virus-induced gene silencing (VIGS), particularly in plant species that are recalcitrant to stable genetic transformation [3] [71].
Within the context of reverse genetics technology, VIGS utilizes recombinant viral vectors to trigger systemic suppression of endogenous plant gene expression, enabling functional characterization through observable phenotypic changes [3]. However, the efficiency of conventional VIGS is often limited by the host's intrinsic RNA silencing machinery, which targets both the viral vector and the endogenous genes it aims to silence. The integration of VSRs addresses this limitation by temporarily inhibiting components of the host's antiviral RNAi pathway, thereby allowing for more robust and widespread silencing of target genes [72] [71]. This technical guide examines the mechanisms of major VSRs and provides detailed protocols for their application in enhancing silencing efficacy for plant functional genomics and recombinant protein expression.
Molecular Mechanisms of Viral Suppressors of RNA Silencing
Key Pathways Targeted by VSRs
VSRs employ diverse strategies to inhibit host RNA silencing, targeting multiple points in the pathway from initiation to amplification. These proteins exhibit remarkable structural diversity with minimal sequence similarity, yet they converge on critical regulatory nodes within the silencing machinery [70]. The most common mechanisms include: (1) dsRNA binding to prevent dicing, (2) siRNA sequestration to inhibit RISC loading, (3) direct interference with AGO protein function, and (4) suppression of secondary siRNA amplification [70]. The diagram below illustrates the primary mechanisms and cellular localization of major VSR activities.
Characterization of Major VSR Classes
dsRNA-Binding VSRs
Proteins such as P38 from Turnip crinkle virus (TCV) and P14 from Pothos latent aureusvirus bind to long dsRNA substrates in a size-independent manner, effectively preventing the processing of viral dsRNA into siRNAs by Dicer-like (DCL) enzymes [70]. This inhibition occurs at the initial step of the RNA silencing pathway, reducing the pool of primary virus-derived siRNAs (vsiRNAs) available for RISC loading.
siRNA-Binding VSRs
The P19 protein from Tombusviruses represents the best-characterized class of VSRs that specifically bind siRNAs. P19 forms a head-to-tail homodimer that measures and sequesters 21-nucleotide siRNA duplexes, preventing their incorporation into RISC [70]. Recent research has revealed that P19 also employs a secondary mechanism through the specific induction of miR168, which post-transcriptionally regulates AGO1 mRNA, thereby reducing AGO1 protein accumulation [70].
AGO-Targeting VSRs
Multiple VSRs directly target ARGONAUTE proteins, the catalytic components of RISC. The C2b protein from Cucumber mosaic virus (CMV) physically interacts with the PAZ and PIWI domains of AGO1, inhibiting its slicing activity [70]. Similarly, the P0 protein from Poleroviruses contains a minimal F-box domain that targets AGO proteins for degradation via the ubiquitin-proteasome pathway [70]. Some VSRs, including the P1 protein from Sweet potato mild mottle virus (SPMMV), mimic host GW/WG repetitive motifs ("AGO hooks") to competitively disrupt the binding of endogenous RNAi components to AGO proteins [70].
VSRs Inhibiting Silencing Amplification
Several VSRs suppress the amplification of antiviral silencing by targeting host RNA-dependent RNA polymerases (RDRs). The V2 protein from Tomato yellow leaf curl virus (TYLCV) binds to SGS3, a cofactor of RDR6, thereby inhibiting the conversion of single-stranded RNA to dsRNA for secondary siRNA production [70]. Similarly, the C2b protein of CMV has been shown to inhibit RDR-dependent secondary vsiRNA formation, although the precise mechanism remains unclear [70].
Experimental Implementation: VSR-Enhanced VIGS Systems
VSR Selection and Vector Engineering
The effective implementation of VSRs to enhance silencing efficacy requires careful selection of appropriate suppressors and their strategic incorporation into viral vectors. Research has demonstrated that different VSRs vary significantly in their potency and specific mechanisms of action, making selection criteria crucial for experimental success [72].
Table 1: Comparison of Major Viral Suppressors of RNA Silencing
| VSR | Viral Source | Mechanism of Action | Key Features | Reported Enhancement |
|---|---|---|---|---|
| P19 | Tombusvirus | Binds and sequesters siRNAs; induces miR168 expression | High-affinity siRNA binding; broad-spectrum suppression | 3-4 fold increase in recombinant protein expression [72] |
| P38 | Turnip crinkle virus | Binds long dsRNA; inhibits DCL4 activity | Size-independent dsRNA binding; suppresses primary siRNA biogenesis | Close second to NSs in protein expression enhancement [72] |
| C2b | Cucumber mosaic virus | Binds dsRNA and AGO1; inhibits RDR-dependent amplification | Dual mechanisms; systemic silencing suppression | Enhanced VIGS efficacy in pepper when truncated (C2bN43) [71] |
| NSs | Tomato spotted wilt virus | Not fully characterized | Potent silencing suppression | Highest recombinant protein expression (0.50 mg/g FW) [72] |
| P0 | Polerovirus | Targets AGO proteins for degradation | F-box mediated ubiquitination | Effective but may cause severe developmental defects |
When engineering viral vectors, the position and orientation of VSR insertion significantly impact performance. Recent studies with Potato virus X (PVX)-derived vectors demonstrated that reversing the VSR cassette orientation relative to the target gene alleviates transcriptional interference, significantly improving both target protein and VSR expression levels [72]. Among the tested configurations, vectors with VSRs inserted in the reverse orientation consistently outperformed conventional designs.
Protocol: Engineering TRV Vectors with Truncated C2b for Enhanced VIGS
Principle
This protocol describes the structure-guided truncation of the CMV C2b protein to decouple its local and systemic silencing suppression activities. The rationale is based on recent findings that selective disruption of local silencing suppression while maintaining systemic suppression enhances TRV-mediated VIGS efficiency in pepper plants [71].
Reagents and Equipment
- pH7lic4.1 expression vector or equivalent
- pTRV2-lic VIGS vector
- cDNA template for CMV C2b
- Restriction enzymes (AgeI, SpeI)
- T4 DNA ligase
- Nicotiana benthamiana and pepper seeds (Capsicum annuum L.)
- Agrobacterium tumefaciens strain GV3101
- LB medium with appropriate antibiotics
- Infiltration buffer (10 mM MES, 10 mM MgCl₂, 200 µM acetosyringone)
Step-by-Step Procedure
Amplification of C2b Truncation Variants
- Design primers to amplify full-length C2b and truncated variants (C2bN43, C2bN69, C2bC79)
- Perform PCR amplification using CMV cDNA as template
- Purify PCR products using gel extraction kit
Cloning into Expression Vector
- Digest both PCR products and pH7lic4.1 vector with AgeI and SpeI restriction enzymes
- Ligate fragments using T4 DNA ligase at 16°C overnight
- Transform into E. coli competent cells and select on LB plates with spectinomycin
- Verify constructs by colony PCR and sequencing
Assembly of TRV-C2bN43 VIGS Vector
- Fuse the C2bN43 fragment with the subgenomic RNA promoter from Pea Early Browning Virus (PEBV) at the 5'-terminus
- Clone the PEBV-C2bN43 cassette into pTRV2-lic vector using Gateway BP/LR clonase or traditional restriction-ligation
- Transform into Agrobacterium GV3101 by electroporation
Agroinfiltration for VIGS
- Inoculate single colonies of Agrobacterium containing pTRV1, pTRV2-C2bN43, and pTRV2-C2bN43-target gene in 5 mL LB medium with appropriate antibiotics
- Shake at 28°C for 24 hours at 200 rpm
- Dilute 1:50 in fresh LB medium with antibiotics and acetosyringone (200 µM)
- Grow to OD₆₀₀ ≈ 0.6-0.8
- Harvest cells by centrifugation and resuspend in infiltration buffer to OD₆₀₀ = 1.0
- Mix pTRV1 and pTRV2-C2bN43-target gene cultures in 1:1 ratio
- Incubate at room temperature for 3-4 hours
- Infiltrate into 2-4 leaf stage pepper plants using needleless syringe
- Maintain inoculated plants at 20°C with 16/8 hour light/dark cycle
Validation and Optimization
- Monitor silencing efficiency by observing phenotypic changes in positive control (e.g., CaPDS silencing causing photobleaching)
- Quantify target gene expression by RT-qPCR at 2-3 weeks post-infiltration
- For anthocyanin-related genes in pepper, assess anther color change as visual marker
- Optimize agroinfiltration density (OD₆₀₀ = 0.3-1.5) for specific plant genotypes
Table 2: Research Reagent Solutions for VSR-Enhanced Silencing
| Reagent/Material | Function/Application | Example Sources | Technical Considerations |
|---|---|---|---|
| pTRV1/pTRV2 Vectors | Bipartite TRV genome for VIGS | Arabidopsis Biological Resource Center | TRV1 encodes replication proteins; TRV2 contains cloning site for target inserts |
| CMV C2b Truncation Mutants | Enhanced VIGS efficiency | Engineered from wild-type C2b [71] | C2bN43 retains systemic suppression while abolishing local suppression |
| pH7lic4.1 Expression Vector | Protein expression and suppressor activity assays | Addgene or similar repositories | CaMV 35S promoter-driven expression with C-terminal 3×Flag tag |
| Agrobacterium GV3101 | Delivery of VIGS constructs into plant cells | Commercial suppliers | Optimal for Solanaceae species; requires acetosyringone for virulence induction |
| pTRV2-C2bN43 Vector | Optimized VIGS in pepper | Custom construction [71] | Specifically enhances silencing in reproductive tissues and difficult-to-silence organs |
Quantitative Assessment of VSR-Enhanced Silencing Efficacy
Performance Metrics and Experimental Outcomes
The efficacy of VSR-enhanced silencing systems must be quantitatively evaluated using both molecular and phenotypic assessments. Recent research provides comprehensive data on the performance improvements achievable through strategic VSR implementation.
Table 3: Quantitative Efficacy of VSR-Enhanced Systems
| Application | VSR System | Performance Metrics | Control Baseline | Enhanced Performance |
|---|---|---|---|---|
| Recombinant Protein Expression | PVX-P19 | GFP accumulation: 0.50 mg/g FW | 0.13 mg/g FW (PVX only) | 3.8-fold increase [72] |
| Vaccine Antigen Production | PVX-NSs | VP1 yield: 0.016 mg/g FW | <0.00016 mg/g FW | >100-fold increase [72] |
| VIGS in Pepper | TRV-C2bN43 | Silencing efficacy in anthers | ~40% with wild-type TRV | ~95% with optimized system [71] |
| Target Gene Knockdown | TRV-C2bN43-CaAN2 | Transcript reduction (RT-qPCR) | No significant reduction | >70% reduction in CaAN2 expression [71] |
The experimental workflow for implementing and validating VSR-enhanced systems involves multiple critical steps from vector construction to quantitative assessment, as illustrated below.
Troubleshooting and Optimization Strategies
Even with optimized VSR systems, researchers may encounter variable silencing efficiency. Common issues and their solutions include:
Incomplete Silencing
- Potential cause: Insfficient VSR activity or incorrect VSR selection
- Solutions: Test multiple VSRs or combinations; optimize agroinfiltration density; use stronger viral vectors
Excessive Viral Symptoms
- Potential cause: Overly potent VSR causing cytotoxicity
- Solutions: Use truncated VSR variants (e.g., C2bN43); reduce incubation temperature; shorten expression time
Tissue-Specific Variations
- Potential cause: Differential VSR activity across tissues
- Solutions: Employ tissue-specific promoters; use vascular-targeted VSRs; optimize plant growth conditions
Host Species-Dependent Efficiency
- Potential cause: Variation in RNA silencing machinery between species
- Solutions: Screen multiple VSRs; consider host-adapted viral vectors; optimize delivery methods
Recent research has demonstrated that environmental factors significantly impact VIGS efficiency. Maintaining plants at 20°C post-inoculation, compared to standard growth temperatures of 25°C, significantly enhances silencing spread and efficacy in pepper plants [71]. Similarly, controlling light intensity (150-200 µE m⁻² s⁻¹) and maintaining consistent humidity (60-70%) improves reproducibility across experiments.
The strategic implementation of viral suppressors of RNA silencing represents a powerful approach to enhance the efficacy of virus-induced gene silencing and recombinant protein expression systems. Through careful selection of appropriate VSRs and optimization of their expression parameters, researchers can achieve substantial improvements in silencing efficiency, particularly in challenging plant species like pepper that have traditionally been recalcitrant to genetic manipulation.
The emerging strategy of functional decoupling through structure-guided truncation, as demonstrated with the C2bN43 mutant, offers a refined approach to VSR engineering that maintains beneficial systemic suppression while minimizing detrimental local effects [71]. This principle of precision engineering of viral proteins is likely to guide future VSR development, potentially leading to designer suppressors with customized properties for specific applications.
As plant biotechnology continues to advance, the integration of VSR-enhanced systems with emerging technologies like CRISPR/Cas9 [73] and multi-omics approaches will further expand our capability to manipulate gene expression in diverse plant species. These integrated approaches will accelerate both basic research in functional genomics and applied biotechnology for crop improvement and recombinant protein production.
VIGS in the Genomics Toolbox: Validation Strategies and Comparative Analysis with CRISPR and RNAi
Virus-induced gene silencing (VIGS) has emerged as a powerful reverse genetics tool for high-throughput functional genomics, enabling researchers to rapidly degrade target gene mRNA without the need for stable transformation [74]. Within this methodology, validation of successful gene silencing stands as a critical component, bridging the gap between experimental execution and biological interpretation. This technical guide examines the integrated validation framework combining phenotypic assessment using visual marker genes, particularly phytoene desaturase (PDS), with molecular confirmation via reverse-transcription quantitative PCR (RT-qPCR). The PDS gene, encoding a key enzyme in the carotenoid biosynthesis pathway, serves as an ideal visual marker because its silencing disrupts chlorophyll production, resulting in easily detectable photobleaching or albino phenotypes across diverse plant species [74] [60] [75]. This dual-approach validation strategy provides researchers with both immediate visual confirmation and precise molecular quantification, ensuring rigorous assessment of VIGS efficacy in functional genomics studies, particularly within the context of plant stress responses, developmental biology, and trait characterization for crop improvement.
PDS as a Visual Marker Gene: Principles and Applications
Biological Function and Silencing Mechanisms
The phytoene desaturase enzyme catalyzes the conversion of phytoene to ζ-carotene in the carotenoid biosynthesis pathway, a crucial step preceding the production of colored carotenoids like lycopene [76]. Silencing of PDS through VIGS or CRISPR/Cas9-mediated knockout disrupts this pathway, leading to photobleaching, albino phenotypes, and dwarfism due to the absence of chlorophyll and carotenoids, which are essential for photosynthesis and photoprotection [74] [75] [76]. This visible phenotype provides an invaluable visual indicator of successful gene silencing without requiring specialized equipment or complex assays. The conserved nature of PDS across plant species makes it particularly useful as a universal marker for method optimization in new plant systems [7] [75].
Beyond its role as a visual marker, PDS silencing has been shown to influence broader physiological processes. Research in tomato fruit demonstrated that PDS silencing not only affects carotenoid biosynthesis genes but also modulates the expression of fruit-ripening genes and ethylene biosynthesis and response genes, suggesting PDS may function as a positive regulator of ripening in tomato fruit [74]. This pleiotropic effect underscores the importance of including appropriate controls and considering potential secondary effects when using PDS as a silencing marker in phenotyping experiments.
Experimental Implementation Across Species
The utility of PDS as a visual marker has been demonstrated across diverse plant species, with optimization of silencing protocols for each system. In soybean, a TRV-based VIGS system utilizing Agrobacterium-mediated infection through cotyledon nodes achieved 65% to 95% silencing efficiency for GmPDS, with photobleaching observed in leaves inoculated with pTRV:GmPDS at 21 days post-inoculation [7]. In highbush blueberry, CRISPR/Cas9-mediated knockout of Vcpds generated albino phenotypes, confirming successful gene editing despite the challenges of polyploid genome editing [75]. Similarly, petunia optimization studies demonstrated that inoculation methods, plant age, temperature, and cultivar selection significantly impact PDS silencing efficiency, with improved protocols increasing PDS silencing by 28% [60].
Table 1: PDS as a Visual Marker in Different Plant Systems
| Plant Species | Validation Method | Key Findings | Silencing Efficiency | Citation |
|---|---|---|---|---|
| Tomato | VIGS (Agroinjection) | Pale-yellow fruit phenotype; affected carotenoid and ripening genes | 100% in two cultivars | [74] |
| Soybean | TRV-VIGS | Photobleaching in leaves at 21 dpi | 65-95% | [7] |
| Highbush Blueberry | CRISPR/Cas9 | Albino phenotypes in edited lines | 2.1-23.8% (varied by gRNA) | [75] |
| Petunia | TRV-VIGS | Optimization of parameters increased silencing | 28% increase with optimization | [60] |
| Cotton | TRV-VIGS | Photobleaching confirmed systemic VIGS | Not specified | [28] |
| Banana | CRISPR/Cas9 & genome identification | PDS markers developed for A/B genome identification | 99.33% accuracy for A genome | [76] |
Molecular Validation: Best Practices in qPCR Analysis
Reference Gene Selection and Validation
Accurate normalization using stable reference genes is fundamental to reliable RT-qPCR analysis, particularly in VIGS experiments where viral infection and gene silencing can alter the expression of commonly used reference genes. A comprehensive study in cotton under VIGS and herbivory stress demonstrated that frequently used reference genes GhUBQ7 and GhUBQ14 were the least stable, while GhACT7 and GhPP2A1 showed the highest stability under these experimental conditions [28]. Similarly, research in arrowhead (Sagittaria trifolia) identified UBQ as the most stable reference gene across multiple tissues, while IDH and PLA were optimal for specific developmental stages [77]. These findings underscore the necessity of empirically validating reference genes for each experimental system rather than relying on conventional choices.
The selection of appropriate reference genes should be guided by statistical algorithms specifically designed for stability analysis. The most commonly employed tools include geNorm, NormFinder, BestKeeper, ΔCt method, and RefFinder, which collectively provide a robust framework for identifying optimal reference genes [77] [28]. For VIGS experiments, it is recommended to use a minimum of two validated reference genes with demonstrated stability under the specific experimental conditions, including the presence of the viral vector and any applied stresses relevant to the research question.
qPCR Experimental Design and Data Analysis
Robust qPCR analysis requires careful experimental design and appropriate statistical approaches to avoid misinterpretation of results. Recent methodological advances highlight limitations of the commonly used 2−ΔΔCT method, which often overlooks variability in amplification efficiency [78]. Instead, Analysis of Covariance (ANCOVA) provides greater statistical power and robustness by modeling raw fluorescence curves and accounting for efficiency variations [78]. This approach enhances rigor and reproducibility, especially when analyzing subtle expression changes.
Adherence to the MIQE (Minimum Information for Publication of Quantitative Real-Time PCR Experiments) guidelines and FAIR (Findable, Accessible, Interoperable, Reproducible) data principles ensures transparent reporting and facilitates data sharing and meta-analyses [78]. Key considerations include providing raw fluorescence data, detailed protocols, primer validation information, and comprehensive analysis scripts. Sharing of qPCR data through general-purpose repositories (e.g., figshare) and code through platforms like GitHub further enhances reproducibility and enables community verification of findings.
Table 2: Key Reagent Solutions for VIGS Validation Experiments
| Reagent/Category | Specific Examples | Function/Application | Technical Considerations |
|---|---|---|---|
| VIGS Vectors | pTRV1, pTRV2 (TRV system) | Viral-induced silencing | pTRV2-empty vector may cause necrosis; pTRV2-sGFP reduces symptoms [60] |
| Agrobacterium Strains | GV3101 | VIGS vector delivery | Resuspended in induction buffer (10 mM MES, 10 mM MgCl2, 200 μM acetosyringone) [74] [7] |
| Visual Marker Constructs | TRV2-PDS | Silencing efficiency visual confirmation | Causes photobleaching; consider pleiotropic effects on pathways of interest [74] |
| Reference Genes | GhACT7, GhPP2A1 (cotton); UBQ (arrowhead) | qPCR normalization | Must be validated for each experimental system; avoid GhUBQ7/GhUBQ14 in cotton-VIGS [28] |
| RNA Extraction Kits | Spectrum Total RNA Kit; Plant Total RNA Kit | RNA isolation | Assess quality (A260/280 ratio 1.8-2.1); purity critical for cDNA synthesis [77] [28] |
| Reverse Transcription Kits | ReverTra Ace-α kit | cDNA synthesis | Use 1μg total RNA with oligo dT20 primer [74] |
| qPCR Master Mixes | iTaq Universal SYBR Green Supermix | Amplification detection | Includes hot-start DNA polymerase, buffer, dNTPs, SYBR Green I dye [77] |
Integrated Experimental Protocols
VIGS Implementation with PDS Marker
The following protocol outlines the optimized VIGS procedure for gene silencing validation, integrating both phenotypic and molecular assessment:
Agrobacterium Preparation and Inoculation:
- Transform A. tumefaciens strain GV3101 with pTRV1 and pTRV2-PDS vectors [7].
- Culture bacteria in LB broth with appropriate antibiotics (25 mg L−1 kanamycin and rifampicin) at 28°C until OD600 reaches 0.6-1.0 [74] [7].
- Centrifuge and resuspend pellets in inoculation buffer (10 mM MgCl2, 10 mM MES pH 5.6, 200 μM acetosyringone) to final OD600 of 1.0 [74].
- Mix pTRV1 and pTRV2-PDS cultures in 1:1 ratio and incubate for 3-6 hours at room temperature [7].
- For tomato fruit: Inject through carpopodium into mature green fruits [74]. For soybean: Use cotyledon node inoculation with 20-30 minute immersion [7]. For petunia: Inoculate mechanically wounded shoot apical meristems [60].
Plant Maintenance and Phenotypic Assessment:
- Maintain inoculated plants in growth chambers at optimized temperatures (20°C day/18°C night for petunia) [60].
- Monitor for photobleaching symptoms beginning at 14-21 days post-inoculation.
- Document silencing frequency using formula: (Number of plants with visible silencing / Total number inoculated) × 100 [74].
- For detached fruit systems, maintain injected fruits at 23°C with 16-h photoperiod and 70% relative humidity [74].
Molecular Validation via RT-qPCR
RNA Extraction and cDNA Synthesis:
- Extract total RNA from silenced tissues (including control tissues) using commercial kits (e.g., Spectrum Total RNA Kit) [28].
- Assess RNA quality and purity (OD260/280 ratio of 1.8-2.1) [77].
- Synthesize cDNA using 1μg total RNA with reverse transcriptase (e.g., ReverTra Ace-α kit) and oligo dT20 primers [74].
qPCR Analysis:
- Design primers with melting temperatures of 54-60°C, GC content 45-55%, and amplicon length of 100-250 bp [77].
- Validate primer efficiency using serial cDNA dilutions (10× to 105×) [77].
- Perform qPCR reactions in technical triplicates using SYBR Green master mixes [77].
- Use the following cycling conditions: pre-denaturation at 95°C for 3 min, followed by 39 cycles of 95°C for 10s and 60°C for 30s [77].
- Include dissociation curve analysis from 65°C to 95°C to verify amplification specificity [77].
Data Analysis:
- Analyze data using ANCOVA or similar robust statistical methods rather than relying solely on 2−ΔΔCT [78].
- Normalize target gene expression using at least two validated reference genes [28].
- Report results following MIQE guidelines, including raw fluorescence data and analysis scripts for reproducibility [78].
Troubleshooting and Technical Considerations
Addressing Common Experimental Challenges
Low Silencing Efficiency: If PDS photobleaching is weak or inconsistent, optimize inoculation methods and plant growth conditions. In petunia, meristem wounding inoculation and younger plants (3-4 weeks) significantly improved silencing efficiency [60]. Temperature optimization is also critical, with 20°C day/18°C night temperatures inducing stronger silencing than higher temperatures in petunia [60].
Viral Symptoms in Controls: Severe necrosis and stunting in empty vector controls can complicate phenotyping. Replacement of pTRV2 empty vector with pTRV2-sGFP containing a fragment of non-plant DNA effectively reduces viral symptoms while maintaining silencing efficiency [60].
Reference Gene Instability: When unexpected reference gene variation occurs, validate additional candidates using multiple statistical algorithms. In cotton-aphid-VIGS studies, GhACT7 and GhPP2A1 demonstrated superior stability compared to traditionally used ubiquitin genes [28].
Species-Specific Optimization: For new plant systems, conduct preliminary experiments with PDS to establish optimal conditions. In soybean, conventional infiltration methods showed low efficiency due to thick cuticles and dense trichomes, while cotyledon node inoculation with extended immersion time significantly improved infection rates [7].
The integration of visual marker genes like PDS with rigorous qPCR validation provides a robust framework for confirming gene silencing in VIGS experiments. This dual-approach validation strategy leverages the immediate feedback of phenotypic markers with the precision of molecular quantification, creating a complementary system that enhances experimental reliability. As VIGS technology continues to evolve, with expanding applications in non-model plants and high-throughput functional genomics, the principles of rigorous validation outlined in this guide will remain fundamental to generating biologically meaningful data.
Future methodological developments will likely focus on multiplexed validation systems, incorporating additional visual markers beyond PDS for simultaneous assessment of multiple genetic elements. Similarly, advances in qPCR methodology, including improved statistical approaches and standardized reporting practices, will further enhance the precision and reproducibility of molecular validation. By adhering to these comprehensive validation protocols, researchers can maximize the utility of VIGS as a powerful reverse genetics tool for elucidating gene function in plant systems.
VIGS Validation Workflow Integration
Virus-induced gene silencing (VIGS) has emerged as a powerful RNA-mediated reverse genetics technology that enables rapid functional genomics studies in plants. This technical guide provides a comprehensive benchmarking analysis of VIGS against stable transformation, focusing on the core parameters of speed, cost, and transient nature within the context of modern plant biology research. As the demand for high-throughput gene function validation accelerates, particularly in non-model species and recalcitrant plants, understanding the precise advantages and limitations of VIGS becomes increasingly critical for researchers and drug development professionals working with plant-derived pharmaceutical compounds [1] [27].
VIGS operates by exploiting the plant's innate RNA interference (RNAi) machinery, using recombinant viruses to carry fragments of target host genes, triggering sequence-specific mRNA degradation and transient gene silencing [1]. Unlike stable transformation, which requires integration of foreign DNA into the plant genome, VIGS functions primarily at the post-transcriptional level without permanent genetic modification [79]. This fundamental distinction underpins the significant differences in experimental timelines, resource requirements, and applications between these two reverse genetics approaches.
Technical Mechanisms and Signaling Pathways
Molecular Mechanism of VIGS
The VIGS process initiates when a recombinant viral vector containing a fragment (typically 200-500 bp) of the plant target gene is introduced into the host plant, most commonly via Agrobacterium tumefaciens-mediated delivery [79] [27]. Once inside plant cells, the viral vector begins replicating and transcribing, producing double-stranded RNA (dsRNA) molecules either through the formation of secondary structures or through the activity of viral RNA-dependent RNA polymerases [1].
These viral dsRNAs are recognized by the plant's Dicer-like (DCL) enzymes, which process them into small interfering RNA (siRNA) duplexes of 21-24 nucleotides in length [1]. The siRNAs are then incorporated into an RNA-induced silencing complex (RISC), where the guide strand directs the complex to complementary endogenous mRNA sequences [1]. The Argonaute (AGO) protein within RISC catalyzes the cleavage and degradation of target mRNAs, resulting in gene silencing [1]. A crucial amplification step occurs when host RNA-dependent RNA polymerases (RDRPs) use the cleaved mRNAs as templates to produce secondary dsRNAs, thereby reinforcing and systemically spreading the silencing signal throughout the plant [1].
Comparative Workflow: VIGS vs. Stable Transformation
The experimental workflows for VIGS and stable transformation differ significantly in both complexity and duration. VIGS procedures typically follow a streamlined path from vector construction to phenotypic analysis, while stable transformation involves multiple additional steps for plant regeneration and selection over extended periods.
Quantitative Benchmarking Analysis
Time Efficiency Comparison
The most significant advantage of VIGS over stable transformation lies in its dramatically reduced experimental timeline. While stable transformation requires multiple months to generate transgenic plants, VIGS can yield functional data within weeks.
Table 1: Time Efficiency Benchmarking
| Experimental Phase | VIGS Timeline | Stable Transformation Timeline | Time Reduction |
|---|---|---|---|
| Vector Construction | 1-2 weeks | 2-4 weeks | ~50% |
| Plant Transformation/Inoculation | 1 day | 1 day | - |
| Selection/Establishment | 1-2 weeks | 2-3 months | ~85-90% |
| Molecular Confirmation | 1 week | 2-4 weeks | ~65-75% |
| Phenotypic Analysis | 1-2 weeks | Ongoing (next generation) | ~90%+ |
| Total Project Duration | 3-6 weeks | 6-12+ months | ~80-90% |
Recent studies demonstrate the remarkable efficiency of optimized VIGS systems. In Camellia drupifera, researchers achieved successful gene silencing within 7-14 days post-inoculation using pericarp cutting immersion techniques, with silencing efficiency reaching approximately 70-91% depending on the target gene and developmental stage [27]. Similarly, in the parasitic plant Striga hermonthica, VIGS-mediated silencing of the phytoene desaturase (PDS) gene produced visible photo-bleaching phenotypes within 7 days via agro-infiltration and 14 days via agro-drench methods [79].
Cost Analysis and Resource Requirements
The transient nature of VIGS translates to substantial cost savings across multiple dimensions, including personnel time, facility space, and consumables.
Table 2: Cost Component Analysis
| Cost Category | VIGS | Stable Transformation | Cost Differential |
|---|---|---|---|
| Personnel Requirements | 2-3 months technical effort | 6-12 months technical effort | 60-75% reduction |
| Facility Space | Limited growth chamber space (weeks) | Extensive greenhouse space (months) | ~70% reduction |
| Consumables | Minimal media and selection agents | Significant media, antibiotics, hormones | ~50% reduction |
| Vector Construction | Standard molecular biology costs | Similar molecular biology costs | Comparable |
| Plant Maintenance | Short-term (weeks) | Long-term (multiple generations) | ~80% reduction |
| Total Project Cost | Low to moderate | High | ~50-70% reduction |
The resource efficiency of VIGS is particularly valuable for high-throughput functional genomics screens, where numerous genes must be tested rapidly with limited budgets. A single VIGS experiment can provide functional data in weeks compared to the many months required for stable transformation, enabling researchers to prioritize candidates for more resource-intensive stable transformation approaches [27].
Key Methodological Protocols
VIGS Implementation in Recalcitrant Species
Recent methodological advances have extended VIGS applications to previously challenging plant species. The following protocol for woody species like Camellia drupifera demonstrates the adaptability of VIGS systems:
Vector Construction: Modified Tobacco rattle virus (TRV) vectors, specifically pNC-TRV2 and pNC-TRV2-GFP, are used. Target gene fragments (200-300 bp) with high specificity are selected using the SGN VIGS Tool and cloned into the TRV2 vector [27].
Agrobacterium Preparation: Recombinant plasmids are transformed into Agrobacterium strains such as GV3101. Cultures are grown in YEB medium with appropriate antibiotics (25 μg/mL kanamycin, 50 μg/mL rifampicin) and induced with acetosyringone (200 μM) when OD₆₀₀ reaches 0.9-1.0 [27].
Inoculation Methods: For recalcitrant tissues, four infiltration approaches are compared:
- Peduncle injection
- Direct pericarp injection
- Pericarp cutting immersion (most efficient at ~94% success rate)
- Fruit-bearing shoot infusion [27]
Developmental Timing: Optimal silencing efficiency varies with developmental stages, achieving 69.80% for CdCRY1 at early stages and 90.91% for CdLAC15 at mid stages of capsule development [27].
Traditional Stable Transformation Protocol
For comparative purposes, standard stable transformation methodology includes:
Vector Construction: Binary vectors containing target genes with appropriate promoters (e.g., 35S), selectable markers (e.g., kanamycin resistance), and reporter genes (e.g., GFP) are constructed.
Plant Transformation: Agrobacterium-mediated transformation or biolistics are used to introduce DNA into plant tissues, followed by co-cultivation.
Selection and Regeneration: Transformed tissues are selected using antibiotics or herbicides and regenerated into whole plants through organogenesis or embryogenesis over 2-3 months.
Molecular Confirmation: PCR, Southern blotting, and expression analyses confirm stable integration and inheritance of transgenes.
Seed Production: T1 seeds are harvested and screened for Mendelian inheritance patterns, requiring an additional generation (2-3 months).
The Scientist's Toolkit: Essential Research Reagents
Table 3: Key Research Reagent Solutions for VIGS
| Reagent/Resource | Function | Examples/Specifications |
|---|---|---|
| Viral Vectors | Delivery of target gene fragments | TRV (Tobacco Rattle Virus), TMV (Tobacco Mosaic Virus), ALSV (Apple Latent Spherical Virus) |
| Agrobacterium Strains | Biological delivery vector | GV3101, LBA4404, EHA105 |
| Selection Antibiotics | Bacterial and plant selection | Kanamycin (25-100 μg/mL), Rifampicin (50 μg/mL) |
| Induction Compounds | Vir gene induction | Acetosyringone (100-200 μM), MES buffer (pH 5.6) |
| Infiltration Media | Plant inoculation | Liquid YEB or LB media, MgCl₂ solution |
| Target Gene Fragments | Silencing specificity | 200-500 bp fragments with <40% similarity to non-target genes |
| Validation Primers | Efficiency confirmation | qRT-PCR primers outside VIGS target region |
Transient Nature: Advantages and Limitations
Epigenetic Applications and Heritable Modifications
While VIGS is primarily transient, recent research has demonstrated its potential for inducing heritable epigenetic modifications through RNA-directed DNA methylation (RdDM). This emerging application bridges the gap between transient silencing and stable epigenetic inheritance:
Mechanism: VIGS can trigger de novo DNA methylation when the viral vector insert corresponds to promoter regions rather than coding sequences. This process involves the canonical PolIV-RdDM pathway, where 24-nt siRNAs direct DNA methyltransferases to specific genomic loci [1].
Heritability: Studies with Arabidopsis thaliana have shown that TRV:FWAᵗʳ infection leads to transgenerational epigenetic silencing of the FWA promoter sequence, with DNA methylation patterns maintained over multiple generations without the viral vector [1].
Applications: VIGS-induced epigenetic modifications enable the development of stable epi-genotypes with desired traits, potentially revolutionizing plant breeding programs without permanent genetic alteration [1].
Technical Limitations and Considerations
Despite its advantages, VIGS presents several important limitations that researchers must consider:
Transient Expression: Silencing effects are temporary, typically lasting 3-8 weeks depending on the plant species and viral vector [79] [27]. In Striga hermonthica, plants recovered from PDS silencing phenotypes within 28 days post-inoculation [79].
Variable Efficiency: Silencing efficiency can vary significantly between tissues (60.2% via agro-infiltration vs. 10.3% via agro-drench in Striga) and developmental stages [79].
Off-Target Effects: Potential for unintended silencing of genes with high sequence similarity requires careful fragment selection and bioinformatic analysis [27].
Species-Specific Optimization: VIGS protocols require significant optimization for different plant species, particularly for recalcitrant woody plants [27].
VIGS represents a transformative RNA-mediated reverse genetics technology that offers substantial advantages in speed and cost-efficiency compared to stable transformation, while its transient nature presents both limitations and unique opportunities for functional genomics research. The dramatically reduced timelines (weeks versus months) and significant cost savings (50-70% reduction) make VIGS particularly valuable for high-throughput gene validation, preliminary screens, and research in recalcitrant species where stable transformation remains challenging. Recent advances in understanding VIGS-induced epigenetic modifications further expand its potential applications beyond transient silencing to include heritable epigenetic engineering. For drug development professionals and plant researchers, VIGS provides a powerful complementary approach to stable transformation, enabling rapid validation of gene function and accelerating the characterization of biosynthetic pathways for pharmaceutical compounds, as demonstrated in recent studies of withanolide biosynthesis in Solanaceae plants [80].
The advancement of functional genomics relies on technologies that enable precise manipulation and analysis of gene function. Within this landscape, Virus-Induced Gene Silencing (VIGS) and CRISPR/Cas systems have emerged as powerful, yet fundamentally distinct, tools for genetic research [3] [81]. While the CRISPR/Cas system functions primarily as a genome-editing tool capable of creating permanent DNA sequence changes, VIGS operates as a RNA-mediated reverse genetics technology that induces transient gene silencing at the post-transcriptional level [3] [1]. Rather than existing in competition, these technologies offer complementary strengths that can be strategically leveraged for different research objectives. This technical guide examines the core mechanisms, applications, and limitations of both VIGS and CRISPR/Cas, with a specific focus on their evolving roles in functional genomics and the emerging frontier of epigenetic editing. The content is framed within the context of a broader thesis on VIGS RNA-mediated reverse genetics technology research, providing scientists and drug development professionals with a contemporary comparison of these indispensable methodologies.
Fundamental Mechanisms and Technological Bases
Virus-Induced Gene Silencing (VIGS)
VIGS is a technology that exploits the plant's innate antiviral RNA interference (RNAi) machinery to achieve transient gene silencing [30]. The process begins when a recombinant viral vector, carrying a fragment of the host target gene, is introduced into the plant typically via agroinfiltration or other inoculation methods [3] [27]. Once inside the plant cell, the viral vector replicates and spreads systemically, producing double-stranded RNA (dsRNA) during its replication cycle [1]. This dsRNA is recognized by the plant's Dicer-like enzymes (DCL), which process it into 21-24 nucleotide small interfering RNAs (siRNAs) [3] [1]. These siRNAs are then incorporated into the RNA-induced silencing complex (RISC), which guides the complex to complementary endogenous mRNA transcripts for sequence-specific cleavage and degradation, thereby silencing the target gene without altering its DNA sequence [3] [1]. This entire process represents Post-Transcriptional Gene Silencing (PTGS) and typically occurs within 2-3 weeks after inoculation [30].
The efficacy of VIGS is influenced by multiple factors, including vector design, the plant's developmental stage, genotype, and environmental conditions such as temperature and light [3]. Recent research has revealed that VIGS can also induce heritable epigenetic modifications through RNA-directed DNA methylation (RdDM), leading to stable transcriptional gene silencing (TGS) that persists across generations even after the viral vector has been cleared [1]. This epigenetic dimension significantly expands VIGS's applications beyond transient knockdowns.
CRISPR/Cas Systems
CRISPR/Cas systems function as precise genome editing tools that create targeted double-strand breaks (DSBs) in DNA [81]. The core mechanism involves a Cas nuclease (most commonly Cas9) complexed with a programmable guide RNA (gRNA) that directs the nuclease to a specific genomic locus through complementary base pairing [82] [81]. The requirement for a Protospacer Adjacent Motif (PAM) sequence adjacent to the target site ensures precise targeting [81]. Cellular repair mechanisms then address these induced breaks primarily through two pathways: error-prone Non-Homologous End Joining (NHEJ), which often results in insertion/deletion mutations (indels) that disrupt gene function, or Homology-Directed Repair (HDR), which can introduce precise genetic modifications when a donor DNA template is provided [81].
The CRISPR toolbox has expanded significantly beyond standard nucleases to include more precise editing tools such as Base Editors (BEs) and Prime Editors (PEs) [81]. Base Editors enable direct chemical conversion of one DNA base to another (C→T or A→G) without creating DSBs, while Prime Editors use a reverse transcriptase fused to a Cas9 nickase to directly write new genetic information into a target site [81]. These advanced editors have proven particularly valuable for modeling and correcting point mutations, which constitute over 50% of known pathogenic human mutations [81].
The diagram below illustrates the fundamental mechanisms of both technologies:
Comparative Analysis of VIGS and CRISPR/Cas Technologies
The strategic selection between VIGS and CRISPR/Cas technologies requires a thorough understanding of their respective capabilities, limitations, and optimal applications. The following table provides a detailed comparison of their core characteristics:
Table 1: Technical Comparison of VIGS and CRISPR/Cas Technologies
| Parameter | Virus-Induced Gene Silencing (VIGS) | CRISPR/Cas Systems |
|---|---|---|
| Primary Mechanism | RNA-mediated post-transcriptional gene silencing (PTGS) [1] [30] | DNA-level genome editing through double-strand breaks or base modification [81] |
| Molecular Basis | Utilizes plant RNAi machinery; involves siRNA production and RISC complex formation [3] [1] | Relies on bacterial adaptive immunity; uses Cas nucleases and guide RNAs [82] [81] |
| Genetic Outcome | Transient gene knockdown (can induce heritable epigenetic modifications) [1] | Permanent genetic modifications (knockout, knock-in, or point mutations) [81] |
| Timeframe | Relatively fast (2-3 weeks for phenotype observation) [30] | Lengthy process (3-6 months for stable lines) [83] |
| Efficiency | Variable (depends on vector, target gene, and host species) [3] | Generally high but cell-type dependent [83] [81] |
| Throughput Capability | Suitable for moderate to high-throughput screening [3] [1] | Excellent for high-throughput screening with pooled gRNA libraries [81] |
| Multiplexing Capacity | Limited | Excellent (multiple gRNAs can be used simultaneously) [81] |
| Delivery Methods | Agroinfiltration, biolistic delivery, spray methods [3] [30] | Viral vectors, lipid nanoparticles, electroporation, microinjection [84] [82] |
| Host Range | Broad (successful in >50 plant species) [3] | Extremely broad (prokaryotes to eukaryotes) [81] |
| Tissue Culture Requirement | Not required [30] | Required for stable transformation [85] |
| Transgene Integration | No integration (transient expression) [85] | Can be integrated or transient |
| Regulatory Status | May face fewer regulatory hurdles in agriculture [85] | Subject to GMO regulations in many countries [85] |
Key Advantages and Limitations
VIGS Advantages and Challenges
VIGS offers several distinctive advantages for functional genomics research. Its transient nature bypasses the need for stable transformation, making it particularly valuable for studying essential genes that might be lethal when permanently disrupted [30]. The methodology is relatively simple, often involving agroinfiltration or similar inoculation techniques, with results typically observable within weeks rather than months [30]. This speed makes VIGS ideal for rapid gene function screening. Furthermore, VIGS is particularly advantageous for plant species that are recalcitrant to transformation, as demonstrated by its successful application in numerous crops including pepper, tomato, and even challenging woody plants like Camellia species [3] [27]. Recent research has expanded VIGS applications beyond transient knockdowns to include the induction of heritable epigenetic modifications through virus-induced transcriptional gene silencing (ViTGS), enabling stable phenotypic changes without DNA sequence alteration [1].
The limitations of VIGS include its variable efficiency, which can be influenced by factors such as plant developmental stage, environmental conditions, and viral vector performance [3]. There is also the potential for non-specific effects due to viral infection symptoms or off-target silencing, though careful vector design and appropriate controls can mitigate these concerns [30]. Additionally, the silencing effect may be incomplete or transient, potentially limiting the observation of phenotypes for genes with low turnover rates or high expression levels [3].
CRISPR/Cas Strengths and Limitations
CRISPR/Cas systems offer unparalleled precision and permanence in genetic manipulation, enabling the creation of stable genetic variants for long-term studies [81]. The technology's modular design—where only the gRNA needs to be modified to change targets—makes it highly adaptable and scalable for high-throughput functional genomics screens [81]. The development of diverse CRISPR tools, including base editors and prime editors, has dramatically expanded its application scope beyond simple gene knockouts to include precise nucleotide conversions and gene regulation capabilities [81].
The primary challenges of CRISPR/Cas systems include the persistent issue of delivery efficiency, particularly for in vivo applications in therapeutic contexts [84] [82]. The potential for off-target effects remains a concern, though improved editor designs and delivery methods have substantially reduced this risk [82] [81]. The technology's dependence on cellular repair mechanisms can lead to variable outcomes, especially for HDR-based approaches which compete with the more error-prone NHEJ pathway [81]. Additionally, the regulatory landscape for CRISPR-edited organisms, particularly in agriculture, remains complex and varies significantly between countries [85].
Experimental Protocols and Workflows
VIGS Implementation Protocol
The successful implementation of VIGS requires careful optimization at each step, from vector selection to phenotypic analysis. The following protocol outlines the key stages for establishing an effective VIGS system:
Vector Selection and Preparation: Choose an appropriate viral vector based on the host plant species. Tobacco Rattle Virus (TRV)-based vectors are widely used for Solanaceous plants like pepper and tomato due to their broad host range and efficient systemic movement [3]. For Camellia species and other recalcitrant plants, modified TRV vectors such as pNC-TRV2 have proven effective [27]. The viral vector should be engineered to include a 200-500 bp fragment of the target gene that exhibits minimal homology to non-target genes to ensure specificity [27].
Insert Design and Cloning: Identify optimal target sequences within the gene of interest using bioinformatics tools such as the SGN VIGS Tool (https://vigs.solgenomics.net/) [27]. Design gene-specific primers with appropriate restriction sites for cloning. Amplify the target fragment from cDNA and clone it into the VIGS vector using standard molecular techniques such as ligation-independent cloning or Gateway recombination [27].
Agrobacterium Preparation and Inoculation: Transform the recombinant VIGS vector into Agrobacterium tumefaciens strains such as GV3101. Culture Agrobacterium cells in YEB medium containing appropriate antibiotics (e.g., kanamycin and rifampicin) and induction agents (e.g., acetosyringone in MES buffer) [27]. Harvest cells when OD600 reaches 0.9-1.0 by centrifugation and resuspend in infiltration medium (10 mM MgCl2, 10 mM MES, 200 μM acetosyringone) to final OD600 of 0.5-2.0 [3] [27].
Plant Inoculation: Select plants at optimal developmental stages—typically 2-4 leaf stage for many species. For tender tissues, use syringe infiltration without needle; for recalcitrant tissues like Camellia capsules, employ specialized methods such as pericarp cutting immersion or fruit-bearing shoot infusion [27]. Maintain inoculated plants under controlled environmental conditions (temperature, humidity, photoperiod) to maximize silencing efficiency [3].
Efficiency Assessment and Phenotypic Analysis: Monitor silencing efficiency using visible markers like photobleaching for PDS silencing or color changes in pigmentation genes [27]. Confirm knockdown at the molecular level through qRT-PCR or RNA gel blot analysis. Document phenotypes 2-4 weeks post-inoculation and conduct further biochemical or physiological assays as required [3] [27].
The workflow diagram below illustrates the key stages of VIGS implementation:
CRISPR/Cas Functional Genomics Workflow
For CRISPR/Cas-based functional genomics, the following protocol outlines key steps for conducting knockout screens and functional validation:
Guide RNA Design and Library Construction: Design gRNAs with high on-target efficiency and minimal off-target potential using computational tools like CRISPRscan or ChopChop. For knockout screens, design 3-6 gRNAs per gene targeting early exons to maximize frameshift probability [81]. For base editing applications, ensure the target base falls within the editing window (typically positions 4-8 for SpCas9) [81]. Synthesize oligonucleotide pools and clone into lentiviral vectors for delivery.
Delivery System Selection: Choose appropriate delivery methods based on cell type. For immortalized cell lines, lentiviral transduction offers high efficiency; for primary cells or in vivo applications, consider lipid nanoparticles (LNPs) or adeno-associated viruses (AAVs) [84] [81]. Recent advances have demonstrated that LNP-mediated delivery enables redosing opportunities not feasible with viral vectors due to immune reactions [84].
Editing and Selection: Transduce cells at appropriate multiplicity of infection (MOI) to ensure single copy integration. For knockout screens, select with appropriate antibiotics (e.g., puromycin) for 3-7 days. For precise edits using HDR, incorporate fluorescence-based sorting or other selection strategies to enrich successfully edited cells [81].
Phenotypic Screening and Analysis: Apply selective pressure relevant to the biological question (e.g., drug treatment, nutrient stress, or cellular stressors). For positive selection screens, identify gRNAs enriched in surviving populations; for negative selection screens, identify depleted gRNAs [81]. Use next-generation sequencing to quantify gRNA abundance and computational tools to identify hits.
Validation and Follow-up: Validate hits using orthogonal approaches, including individual gRNA validation, Western blotting to confirm protein loss, and functional assays. For disease modeling, establish isogenic cell lines with specific mutations to confirm phenotype-genotype relationships [81].
Applications in Functional Genomics and Epigenetic Editing
VIGS Applications in Plant Functional Genomics
VIGS has become an indispensable tool for plant functional genomics, particularly for species recalcitrant to stable transformation. In pepper (Capsicum annuum L.), VIGS has successfully identified genes governing critical agronomic traits including fruit quality (color, biochemical composition, pungency), disease resistance (to bacteria, oomycetes, insects), and abiotic stress tolerance (temperature, salt, osmotic stress) [3]. The technology has been particularly valuable for characterizing genes involved in specialized metabolism, such as the capsaicinoid biosynthesis pathway, where stable genetic transformation remains challenging [3].
Recent applications have extended VIGS to epigenetic studies, where viral vectors carrying sequences homologous to gene promoters rather than coding sequences can induce RNA-directed DNA methylation (RdDM) and transcriptional gene silencing [1]. This approach, known as virus-induced transcriptional gene silencing (ViTGS), has been used to achieve transgenerational epigenetic silencing of the FWA gene in Arabidopsis, with DNA methylation patterns maintained over multiple generations even after the viral vector was eliminated [1]. This application demonstrates VIGS's potential for creating stable epigenetic variation for breeding programs without introducing permanent genetic changes.
CRISPR/Cas in Functional Genomics and Disease Modeling
CRISPR/Cas systems have revolutionized functional genomics by enabling systematic genome-wide knockout screens in diverse cell types and organisms. Pooled CRISPR screens have identified essential genes, drug resistance mechanisms, and novel therapeutic targets across various disease contexts [81]. The technology's precision has facilitated the creation of isogenic cell models that differ only at specific loci of interest, allowing researchers to dissect the functional consequences of individual mutations with unprecedented accuracy [81].
In therapeutic development, CRISPR/Cas has shown remarkable success in clinical trials for genetic disorders. The first FDA-approved CRISPR therapy, Casgevy, treats sickle cell disease and transfusion-dependent beta thalassemia by editing the BCL11A gene to reactivate fetal hemoglobin production [84]. Ongoing clinical trials are exploring CRISPR-based treatments for hereditary transthyretin amyloidosis (hATTR), hereditary angioedema (HAE), and various cancers [84]. Recent breakthroughs include the first personalized in vivo CRISPR treatment for an infant with CPS1 deficiency, developed and delivered in just six months—demonstrating the technology's potential for addressing rare genetic diseases [84].
Emerging Integration of VIGS and CRISPR Technologies
The convergence of VIGS and CRISPR technologies represents an exciting frontier in genetic manipulation. Virus-Induced Genome Editing (VIGE) leverages viral vectors to deliver CRISPR components into plant cells, combining the high efficiency and broad tissue coverage of viral vectors with the precision of CRISPR editing [85]. This approach potentially enables the generation of transgene-free edited plants in a single generation without the need for tissue culture, addressing one of the major bottlenecks in plant genome editing [85]. VIGE has been successfully applied in more than 14 plant species using over 20 different viruses, though challenges remain regarding vector capacity, host immune responses, and editing efficiency in meristematic tissues [85].
The Scientist's Toolkit: Essential Research Reagents and Materials
Successful implementation of VIGS and CRISPR/Cas technologies requires specific reagents and materials optimized for each system. The following table details essential components for both platforms:
Table 2: Essential Research Reagents and Materials for VIGS and CRISPR/Cas Studies
| Category | Specific Reagents/Materials | Function/Application | Examples/Sources |
|---|---|---|---|
| VIGS Vectors | TRV-based vectors (pTRV1, pTRV2) | Bipartite viral vector system for Solanaceous plants [3] | Modified versions (pNC-TRV2) for recalcitrant species [27] |
| BBWV2, CMV, Geminivirus vectors | Alternative vectors for specific host plants [3] | Bean Yellow Dwarf Virus for monocots [3] | |
| CRISPR Editors | Cas9 Nucleases | Creates double-strand breaks for gene knockout [81] | SpCas9, SaCas9 (different PAM requirements) [81] |
| Base Editors (BE) | Enables precise single nucleotide changes without DSBs [81] | ABE (A→G), CBE (C→T), CGBE (C→G) [81] | |
| Prime Editors (PE) | Inserts targeted small indels and all base-to-base conversions [81] | PE2, PE3 systems with engineered reverse transcriptase [81] | |
| Delivery Tools | Agrobacterium tumefaciens | Standard for plant VIGS delivery [3] [27] | GV3101, LBA4404 strains with appropriate virulence |
| Lipid Nanoparticles (LNPs) | In vivo delivery of CRISPR components [84] | Enable redosing without immune reactions [84] | |
| Lentiviral Vectors | High-efficiency delivery for cell culture screens [81] | Used for pooled CRISPR library delivery [81] | |
| Selection Agents | Antibiotics | Selection of transformed bacteria/plant tissues [27] | Kanamycin, rifampicin for Agrobacterium [27] |
| Fluorescent Markers | Visual tracking of transformation/editing efficiency [27] | GFP, RFP in vector systems [27] | |
| Analysis Tools | SGN VIGS Tool | Bioinformatics for optimal target sequence selection [27] | https://vigs.solgenomics.net/ [27] |
| ICE Analysis | Computational analysis of CRISPR editing efficiency [83] | Inference of CRISPR Edits from Sanger sequencing [83] | |
| Specialized Kits | RNAprep Pure Kit | High-quality RNA extraction for silencing verification [27] | Tiangen, China [27] |
| Hieff PCR Master Mix | High-fidelity amplification for vector construction [27] | Yeasen, China [27] |
VIGS and CRISPR/Cas technologies represent complementary pillars of modern functional genomics, each with distinct advantages and optimal applications. VIGS excels as a rapid, cost-effective tool for transient gene silencing and epigenetic studies, particularly in plant species recalcitrant to stable transformation. Its ability to induce heritable epigenetic modifications through RdDM expands its utility beyond transient knockdowns to include stable phenotypic alterations. CRISPR/Cas systems offer unparalleled precision for permanent genetic modifications, enabling the creation of stable knockout lines, precise nucleotide conversions, and sophisticated disease models. The integration of these technologies through approaches like VIGE represents the next frontier in genetic manipulation, combining the efficiency of viral delivery with the precision of CRISPR editing. As both technologies continue to evolve, their strategic application will accelerate functional genomics research and therapeutic development across diverse biological systems.
Virus-induced gene silencing (VIGS) has established itself as a cornerstone RNA-mediated reverse genetics technology in plant research, allowing for rapid functional analysis of genes by exploiting the plant's innate antiviral defense mechanisms [1] [3]. This powerful technique leverages recombinant viral vectors to trigger sequence-specific degradation of target mRNA through the post-transcriptional gene silencing (PTGS) pathway, leading to transient knockdown of gene expression without the need for stable transformation [1] [86]. The significance of VIGS lies in its speed, cost-effectiveness, and applicability to a wide range of plant species, including those recalcitrant to stable genetic transformation [3] [27].
However, VIGS operates within a broader landscape of gene silencing technologies. A comprehensive understanding of this landscape is crucial for researchers to select the most appropriate tool for their specific experimental needs. This review provides a detailed comparative analysis of VIGS against three other prominent silencing methodologies: Host-Induced Gene Silencing (HIGS), Spray-Induced Gene Silencing (SIGS), and Transcription Activator-Like Effector-based (TALE-based) Repressors. By examining the molecular mechanisms, experimental protocols, applications, and relative advantages of each system, this analysis aims to serve as a technical guide for scientists navigating the complex toolkit of modern plant functional genomics and biotechnology.
Virus-Induced Gene Silencing (VIGS)
The VIGS process initiates when a recombinant viral vector, carrying a fragment of the host plant's target gene, is introduced into the plant via agroinfiltration or other inoculation methods [3] [86]. Once inside the plant cell, the viral genome is uncoated and replicates. The plant's antiviral defense machinery recognizes the viral double-stranded RNA (dsRNA) replication intermediates. These dsRNAs are cleaved by the host's Dicer-like (DCL) enzymes into 21-24 nucleotide small interfering RNAs (siRNAs) [1] [86]. These siRNAs are then incorporated into the RNA-induced silencing complex (RISC), where the Argonaute (AGO) protein uses the siRNA as a guide to identify and catalyze the sequence-specific cleavage of complementary endogenous mRNA transcripts, thereby silencing the target gene [1] [86]. A key feature of VIGS is the amplification of the silencing signal by host RNA-directed RNA polymerases (RDRs), which can generate secondary siRNAs, leading to systemic spread of the silencing effect throughout the plant [1]. Furthermore, VIGS can induce heritable epigenetic modifications through RNA-directed DNA methylation (RdDM), leading to transcriptional gene silencing (TGS) when the viral insert targets promoter sequences [1].
Figure 1: Molecular Mechanism of VIGS. The process begins with the introduction of a recombinant viral vector, leading to the production of dsRNA, which is processed into siRNAs. These siRNAs are loaded into RISC, leading to target mRNA cleavage. Additionally, siRNAs can trigger epigenetic modifications via the RdDM pathway.
Host-Induced Gene Silencing (HIGS)
HIGS is a powerful RNAi-based approach that involves engineering a host plant to express dsRNA or artificial microRNAs (amiRNAs) that target essential genes in attacking pathogens or pests [86]. The silencing signal is produced in the plant and then taken up by the pest or pathogen during infection or feeding, triggering RNAi within the attacker's cells and compromising its viability [86]. A landmark study demonstrated this by expressing dsRNA targeting the root-knot nematode parasitism gene 16D10 in Arabidopsis plants, which conferred effective resistance against multiple nematode species [86].
Spray-Induced Gene Silencing (SIGS)
SIGS represents a topical application-based RNAi strategy. Also known as exogenous RNAi (exo-RNAi), it involves the direct application of dsRNAs, siRNAs, or short hairpin RNAs (shRNAs) onto plant surfaces [86]. These externally applied RNA molecules are taken up by the plant cells and/or the pathogen/pest, initiating gene silencing. Delivery methods for SIGS include spraying, infiltration, and root or seed soaking [86]. This method offers a flexible and non-transgenic approach to crop protection.
TALE-based Repressors
Unlike the RNAi-based mechanisms of VIGS, HIGS, and SIGS, TALE-based repressors operate at the transcriptional level. This technology involves the design of artificial Transcription Activator-Like Effectors (TALEs) fused to potent repressor domains [86]. The DNA-binding domain of the TALE is engineered to recognize a specific promoter sequence of the target gene. Upon binding, the fused repressor domain, such as the EAR repression domain (SRDX) in plants, recruits chromatin-modifying complexes that establish a repressive chromatin state, leading to reduced transcription initiation [86]. This results in direct transcriptional silencing of the target gene.
Comparative Analysis of Silencing Technologies
The table below provides a systematic comparison of the key technical and application parameters of VIGS, HIGS, SIGS, and TALE-based Repressors.
Table 1: Comparative Analysis of Gene Silencing Technologies
| Feature | VIGS | HIGS | SIGS | TALE-based Repressors |
|---|---|---|---|---|
| Mechanism of Action | Post-transcriptional gene silencing (PTGS) & RNA-directed DNA methylation (RdDM) [1] [86] | RNAi within the pathogen/pest via in-planta produced dsRNA [86] | RNAi via exogenously applied RNA molecules [86] | Transcriptional gene silencing (TGS) via engineered DNA-binding repressors [86] |
| Primary Application | Rapid functional genomics in plants; gene characterization [1] [3] | Crop protection against pathogens and pests (e.g., nematodes, fungi) [86] | Crop protection; non-transgenic pest/pathogen control [86] | Targeted gene repression in plant and animal systems; functional genomics [86] |
| Duration of Effect | Transient (weeks to months) [3] | Sustained (as long as the transgene is expressed) | Transient (days to weeks) [86] | Stable (in transgenic lines) |
| Organism/Tissue Specificity | Plant-specific; can be tissue-specific depending on the viral vector [3] [27] | Cross-kingdom; targets specific pests/pathogens interacting with the host | Cross-kingdom; targets specific pests/pathogens; limited by uptake and stability [86] | Broad eukaryote applicability; highly sequence-specific within the host genome [86] |
| Key Advantages | Rapid, does not require stable transformation, applicable to recalcitrant species, heritable epigenetic modifications possible [1] [27] | Pesticide-free, highly specific to the target pest/pathogen, durable resistance [86] | Non-transgenic, flexible application, suitable for topical pest management [86] | Precise transcriptional control, high specificity, stable inheritance in transgenic lines [86] |
| Key Limitations | Off-target effects, variable efficiency, influenced by environment, viral symptoms may occur [3] [86] | Requires stable transformation, potential for off-target effects in host, regulatory hurdles for GMOs [86] | RNA molecule stability, environmental degradation, variable uptake efficiency [86] | Complex protein engineering, potential cytotoxicity, off-target binding possible [86] |
Detailed Experimental Protocols
VIGS Protocol for Recalcitrant Plant Tissues
Optimizing VIGS for challenging plant materials, such as the lignified capsules of Camellia drupifera, requires methodical optimization [27]. The following protocol, proven to achieve >90% infiltration efficiency, outlines the critical steps.
Step 1: Vector Construction and Agrobacterium Preparation
- Clone Target Fragment: Amplify a 200-300 bp specific fragment of the target gene (e.g., CdCRY1 or CdLAC15). Screen the fragment for specificity using tools like the SGN VIGS Tool to ensure <40% similarity to non-target genes [27].
- Gateway Cloning: Ligate the fragment into a TRV2-based vector (e.g., pNC-TRV2) using Nimble Cloning or Gateway recombination. Verify the construct by sequencing [27].
- Transform Agrobacterium: Introduce the recombinant TRV2 and the helper TRV1 plasmids into Agrobacterium tumefaciens strains like GV3101.
- Culture Agrobacteria: Grow single colonies in YEB medium supplemented with kanamycin (25 μg/mL) and rifampicin (50 μg/mL) at 28°C for 48 hours. Subculture into a fresh, larger volume of induction medium (with 10 mM MES and 200 μM acetosyringone, pH 5.6) and grow until OD₆₀₀ reaches 0.9-1.0 [27].
- Prepare Inoculum: Centrifuge the bacterial culture and resuspend the pellet in an infiltration buffer (10 mM MgCl₂, 10 mM MES, 200 μM acetosyringone). Adjust the final OD₆₀₀ to a pre-optimized concentration (e.g., 1.5 for C. drupifera). Incubate the mixture for 3-4 hours at room temperature before inoculation [27].
Step 2: Plant Inoculation
- Select Plant Material: Identify the optimal developmental stage for inoculation. For C. drupifera capsules, the highest silencing efficiency for CdLAC15 (~90.91%) was achieved at the mid-stage of development [27].
- Inoculation Method - Pericarp Cutting Immersion: For firm fruits, the most effective method is to make shallow cuts on the pericarp and immediately immerse the fruit in the Agrobacterium inoculum for 15 minutes, applying a mild vacuum if necessary to enhance infiltration [27].
- Control Groups: Always include negative controls (plants inoculated with TRV1 + empty TRV2 vector) and positive controls (plants inoculated with TRV1 + TRV2 carrying a marker gene like PDS).
Step 3: Post-Inoculation Care and Analysis
- Plant Maintenance: Maintain inoculated plants in a greenhouse under controlled conditions. For many species, lowering the temperature to 20-22°C post-inoculation can enhance VIGS efficiency by reducing viral replication speed and mitigating symptoms [86] [6].
- Phenotypic Monitoring: Observe and document the emergence of silencing phenotypes (e.g., photobleaching for PDS, color fading for pigment genes) over 2-6 weeks post-inoculation.
- Molecular Validation: Quantify silencing efficiency by extracting total RNA from target tissues and performing RT-qPCR analysis of the target gene transcript levels, using a stable reference gene (e.g., GAPDH, ACTIN) for normalization [27] [6].
Advanced VIGS Optimization: Engineering Viral Suppressors
To overcome low VIGS efficiency, particularly in reproductive organs, advanced vector engineering can be employed. A recent study demonstrated that a truncated version of the Cucumber mosaic virus 2b (C2b) silencing suppressor, C2bN43, enhances VIGS in pepper [6].
Protocol:
- Generate Truncated VSR: Amplify the N-terminal 43 amino acid fragment of the C2b protein (C2bN43) by PCR.
- Clone into TRV Vector: Fuse the C2bN43 fragment downstream of a subgenomic RNA promoter (e.g., from Pea Early Browning Virus) and clone it into the pTRV2 vector to create pTRV2-C2bN43.
- Assemble Silencing Construct: Insert the target gene fragment (e.g., a 250-bp fragment of CaAN2) into the pTRV2-C2bN43 vector.
- Inoculate and Analyze: Proceed with standard VIGS agroinfiltration protocols. This engineered system retains systemic silencing suppression to promote viral spread but abrogates local suppression, thereby potentiating the RNAi machinery in systemically infected tissues and resulting in significantly stronger silencing [6].
TALE-based Repressor Assembly Protocol
The construction of functional TALE-based repressors involves a multi-step process of protein engineering [86].
Protocol:
- Target Sequence Selection: Identify a unique 14-20 bp DNA sequence within the promoter region of the target gene.
- TALE Repeat Array Assembly: Use modular cloning methods (e.g., Golden Gate assembly) to assemble the TALE DNA-binding domain. The code linking the amino acids at positions 12 and 13 (Repeat Variable Diresidue, RVD) in each TALE repeat dictates nucleotide specificity (e.g., NI for A, NG for T, HD for C, NN for G).
- Repressor Domain Fusion: Fuse the assembled TALE DNA-binding domain to a potent repressor domain. For plants, the most effective domain is often the EAR repression domain (SRDX; 12-aa sequence: LDLDLELRLGFA) [86].
- Vector Construction and Transformation: Clone the final TALE-SRDX fusion construct into a plant binary expression vector under a constitutive promoter (e.g., CaMV 35S). Introduce the vector into plants via Agrobacterium-mediated stable transformation.
- Phenotypic and Molecular Screening: Screen transgenic lines for phenotypic changes and validate transcriptional repression of the target gene using RT-qPCR.
The Scientist's Toolkit: Essential Research Reagents
Table 2: Key Reagent Solutions for Gene Silencing Research
| Reagent / Material | Function / Application | Examples & Technical Notes |
|---|---|---|
| Viral Vectors | Delivery of target sequences to trigger silencing in VIGS. | TRV (Tobacco Rattle Virus): Bipartite system (TRV1/TRV2), broad host range, mild symptoms [3].CMV (Cucumber Mosaic Virus): Can be engineered with suppressors like C2bN43 for enhanced efficiency [6]. |
| Agrobacterium tumefaciens | Biological vector for delivering T-DNA containing viral genomes or silencing constructs into plant cells. | Strain GV3101: Commonly used for agroinfiltration in VIGS and stable transformation for HIGS/TALE-repressors [27]. |
| Induction Agents | Activate the Agrobacterium Vir genes for efficient T-DNA transfer. | Acetosyringone (200 μM): Added to the bacterial culture and infiltration buffer [27].MES buffer (10 mM, pH 5.6): Maintains optimal pH for Vir gene induction. |
| Reporter Genes | Visual markers for assessing silencing efficiency and pattern. | Phytoene Desaturase (PDS): Silencing causes photobleaching, a universal positive control [3] [86].Anthocyanin-related genes (e.g., CaAN2): Silencing disrupts pigmentation in flowers/fruits [6]. |
| Repressor Domains | Effector domains for transcriptional repression in TALE-based systems. | EAR motif (SRDX): The 12-aa sequence (LDLDLELRLGFA) is a potent plant-specific repressor [86].KRAB domain: A common repressor domain used in animal systems, can be tested in plants [86]. |
| High-Fidelity Polymerase | Accurate amplification of target gene fragments for cloning into vectors. | Essential for generating error-free inserts for VIGS, HIGS, and TALE assembly to ensure specificity [27]. |
| Silencing Suppressors (VSRs) | Engineered to enhance VIGS spread and efficiency. | Truncated C2b (C2bN43): Retains systemic but not local suppression, boosting VIGS efficacy in pepper [6]. |
The choice of a gene silencing technology is dictated by the specific research goal. VIGS remains the premier tool for high-throughput, transient functional genomics in plants, especially for species resistant to stable transformation. Its ongoing optimization, for instance through engineered viral suppressors, continues to expand its utility into recalcitrant tissues and organs [27] [6]. HIGS offers a transgenic but highly specific strategy for crop protection, while SIGS provides a complementary, non-transgenic alternative for topical applications. TALE-based Repressors offer a distinct and powerful mechanism for achieving precise transcriptional silencing, filling a critical niche for studies where modulating transcription is required.
The future of gene silencing lies in the strategic combination of these technologies. For example, VIGS can be used for rapid gene validation prior to the development of stable HIGS or TALE-repressor lines. As the molecular mechanisms underlying these systems are further elucidated, and as delivery methods improve, the power, precision, and applicability of this toolkit will continue to grow, profoundly impacting both basic plant research and applied agricultural biotechnology.
Virus-induced gene silencing (VIGS) has emerged as a powerful reverse genetics tool for rapid functional analysis of plant genes. This technology leverages the plant's innate RNA-mediated antiviral defense mechanism, specifically post-transcriptional gene silencing (PTGS), to target and downregulate endogenous genes [1] [5]. The application of VIGS in soybean (Glycine max L.) is particularly valuable for validating disease resistance genes, as stable genetic transformation in this crop remains time-consuming and labor-intensive [7] [87]. This case study focuses on establishing and optimizing a Tobacco Rattle Virus (TRV)-based VIGS system for the functional characterization of key disease resistance genes in soybean, providing a methodological framework for rapid assessment of candidate genes in plant defense mechanisms.
Theoretical Foundations of VIGS
Molecular Mechanisms of Virus-Induced Gene Silencing
VIGS operates through a sophisticated RNA interference pathway that begins when a recombinant virus, carrying a fragment of the target plant gene, is introduced into the host plant [1]. The molecular process unfolds through several key stages:
- Viral Replication and dsRNA Formation: Once inside plant cells, the viral vector replicates, producing double-stranded RNA (dsRNA) intermediates through the action of viral RNA-dependent RNA polymerase (RdRp) [5].
- Dicer-Mediated Processing: The plant's defense system recognizes these dsRNA molecules as foreign. Dicer-like (DCL) enzymes, primarily DCL2 and DCL4, cleave the long dsRNAs into small interfering RNAs (siRNAs) of 21–24 nucleotides in length [1] [5].
- RISC Assembly and Target Degradation: These siRNAs are incorporated into the RNA-induced silencing complex (RISC), where the guide strand directs the complex to complementary endogenous mRNA transcripts. Argonaute (AGO) proteins, the catalytic components of RISC, then cleave the target mRNAs, preventing their translation [1].
- Amplification and Systemic Spread: The silencing signal is amplified by host RNA-dependent RNA polymerases and can spread systemically throughout the plant, leading to a visible phenotype in tissues distant from the initial infection site [5].
Table 1: Key Components of the Plant RNA Silencing Machinery Utilized in VIGS
| Component | Function in VIGS Pathway |
|---|---|
| Dicer-like (DCL) Enzymes | Cleaves viral dsRNA replication intermediates into 21-24 nt siRNAs |
| Small Interfering RNAs (siRNAs) | Guide sequence-specific recognition and degradation of target mRNAs |
| Argonaute (AGO) Proteins | Catalytic component of RISC that slices target mRNAs |
| RNA-dependent RNA Polymerases (RDRs) | Amplifies silencing signal by synthesizing secondary dsRNAs |
The TRV vector is particularly advantageous for VIGS studies due to its ability to infect meristematic tissues, induce mild symptom development, and spread efficiently throughout the plant [5]. Furthermore, VIGS can induce epigenetic modifications in plants, including DNA methylation, which may lead to heritable epigenetic changes, offering potential for long-term trait stability in breeding programs [1].
TRV-VIGS Methodology for Soybean
Vector Construction and Design Principles
The TRV-based VIGS system utilizes a bipartite vector system consisting of two plasmid constructs:
- TRV1 (pTRV1): Encodes viral replication proteins (134K and 194K replicases), a movement protein (MP) for cell-to-cell movement, and a 16K cysteine-rich protein that functions as a weak suppressor of RNA silencing [3] [5].
- TRV2 (pTRV2): Contains the coat protein (CP) gene and a multiple cloning site (MCS) for insertion of target gene fragments [7] [5].
For effective silencing, a 300-500 base pair fragment of the target gene is cloned into the TRV2 vector in sense or antisense orientation [5]. The insert should be designed to avoid highly conserved domains shared with other gene family members unless silencing of multiple genes is intended. Online tools such as the SGN-VIGS tool (https://vigs.solgenomics.net/) can assist in selecting optimal target regions with minimal off-target potential [88].
Table 2: Target Genes and Insert Sizes for Soybean TRV-VIGS
| Target Gene | Function | Insert Size | Phenotypic Outcome of Silencing |
|---|---|---|---|
| GmPDS | Phytoene desaturase in carotenoid biosynthesis | 300-500 bp | Photobleaching (white leaves) |
| GmRpp6907 | Confers resistance to soybean rust | ~400 bp | Compromised rust immunity |
| GmRPT4 | Defense-related gene | ~400 bp | Altered disease response |
| GmPOD53L | Class III peroxidase enhancing lignin deposition | Not specified | Reduced soybean cyst nematode resistance |
Agroinoculation Protocol for Soybean
Soybean's thick leaf cuticle and dense trichomes present challenges for conventional infiltration methods. The optimized protocol involves Agrobacterium-mediated infection through the cotyledon node [7]:
- Plant Material Preparation: Surface-sterilize soybean seeds and germinate on moist vermiculite until radicles reach 1-3 cm in length [7] [88].
- Agrobacterium Preparation: Transform binary TRV vectors into Agrobacterium tumefaciens strain GV3101. Grow bacterial cultures in YEP medium with appropriate antibiotics (50 mg/L kanamycin, 50 mg/L rifampicin) to mid-log phase (OD600 = 0.6-0.8) [7] [88].
- Induction and Resuspension: Pellet bacteria and resuspend in infiltration buffer (10 mM MES, 200 μM acetosyringone, 10 mM MgCl2, 0.03% Silwet-77) to a final OD600 of 0.8-1.0 [88]. Incubate the mixed Agrobacterium suspension (TRV1 + recombinant TRV2) at room temperature for 3 hours in darkness to induce virulence gene expression [88].
- Inoculation: Bisect germinated seeds longitudinally to create half-seed explants. Immerse fresh explants in the Agrobacterium suspension for 20-30 minutes with gentle agitation [7]. For enhanced efficiency, vacuum infiltration (0.5 kPa, 5-10 minutes) can be applied [88].
- Plant Growth and Monitoring: Transfer inoculated plants to vermiculite or nutrient soil and maintain under controlled conditions (22-27°C, 16-hour photoperiod). Silencing phenotypes typically appear in systemic leaves within 14-21 days post-inoculation (dpi) [7].
Diagram 1: TRV-VIGS Experimental Workflow for Soybean (Title: Soybean VIGS Workflow)
Efficiency Optimization and Validation
Successful implementation of TRV-VIGS requires careful optimization and validation:
- Efficiency Assessment: The TRV-VIGS system in soybean demonstrates silencing efficiencies ranging from 65% to 95%, as confirmed by quantitative real-time PCR (qRT-PCR) [7]. Infection efficiency can be monitored using a TRV2-GFP vector, with fluorescence microscopy revealing successful infection in over 80% of cells in the cotyledon node [7].
- Reference Genes for qRT-PCR: For accurate quantification of silencing efficiency, appropriate reference genes must be used. Elongation factor-1α (EF-1) and ubiquitin (ubi3) have been identified as stable reference transcripts in VIGS studies, whereas actin expression may vary significantly under experimental conditions [89].
- Phenotypic Validation: Silencing of the phytoene desaturase (GmPDS) gene serves as a visual marker for VIGS efficiency, resulting in characteristic photobleaching due to chlorophyll degradation [7] [5]. This phenotype typically appears 21 days post-inoculation, initially in cluster buds before spreading to systemic leaves [7].
Validation of Disease Resistance Genes
Case Study 1: Silencing Soybean Rust Resistance Gene GmRpp6907
The TRV-VIGS system was successfully employed to validate the function of GmRpp6907, a gene conferring resistance to soybean rust. Soybean plants silenced for GmRpp6907 showed compromised rust immunity compared to controls, confirming its essential role in disease resistance [7]. This study demonstrated the utility of VIGS for rapid assessment of candidate resistance genes identified through genetic mapping or transcriptomic analyses.
Case Study 2: Functional Analysis of Defense-Related Gene GmRPT4
Silencing of the defense-related gene GmRPT4 using the TRV-VIGS system resulted in significant phenotypic alterations, though the specific nature of these changes was not detailed in the available literature [7]. This case nevertheless illustrates the application of VIGS for characterizing genes involved in plant defense signaling pathways.
Case Study 3: Role of GmPOD53L in Nematode Resistance
In a separate study, TRV-VIGS was used to silence GmPOD53L, a gene encoding a class III peroxidase enzyme. GmPOD53L is upregulated under soybean cyst nematode (SCN) stress and is associated with increased peroxidase activity and lignin deposition [90]. Silencing of GmPOD53L reduced soybean resistance to SCN, demonstrating its positive role in the stress response mechanism. This peroxidase gene enhances lignin content in roots, creating a physical barrier that inhibits nematode invasion and development [90].
Diagram 2: VIGS Molecular Mechanism (Title: VIGS Gene Silencing Mechanism)
The Scientist's Toolkit: Research Reagent Solutions
Table 3: Essential Research Reagents for TRV-VIGS in Soybean
| Reagent/Resource | Function/Application | Specifications/Alternatives |
|---|---|---|
| pTRV1 and pTRV2 Vectors | Binary TRV vectors for VIGS | TRV1 encodes replication and movement proteins; TRV2 with MCS for target insert [5] |
| pTRV2-GFP | Visual tracking of infection | TRV2 vector with GFP reporter for monitoring viral spread [7] |
| Agrobacterium tumefaciens GV3101 | Delivery of TRV vectors into plant cells | Superior for dicot transformation; other strains (e.g., LBA4404) possible [7] [88] |
| Infiltration Buffer | Facilitates Agrobacterium entry | 10 mM MES, 200 μM acetosyringone, 10 mM MgCl₂, 0.03% Silwet-77 [88] |
| SGN-VIGS Tool | Bioinformatics for insert design | Online tool for selecting optimal gene-specific fragments [88] |
| Reference Genes | qRT-PCR normalization | EF-1α and ubiquitin (ubi3) recommended; avoid actin due to variability [89] |
The TRV-VIGS system represents a robust and efficient platform for rapid functional validation of disease resistance genes in soybean. This case study demonstrates its successful application in silencing key genes involved in defense against pathogens such as soybean rust and cyst nematode. The methodology offers significant advantages over stable transformation, including reduced experimental timeline, applicability across diverse soybean genotypes, and the potential for high-throughput screening. Future directions for VIGS technology in soybean research include integration with CRISPR-based systems for enhanced targeting specificity, exploration of VIGS-induced epigenetic modifications for breeding applications, and adaptation for field-scale functional genomics studies. As optimization continues, TRV-VIGS is poised to become an indispensable tool for accelerating the development of disease-resistant soybean cultivars.
Conclusion
Virus-induced gene silencing has firmly established itself as an indispensable, rapid, and versatile tool in the reverse genetics arsenal. By leveraging the plant's own RNAi machinery, VIGS provides unparalleled advantages for high-throughput functional gene validation, particularly in non-model and recalcitrant species where stable transformation is challenging. The technology's evolution, from simple gene knockdown to inducing heritable epigenetic modifications and its integration with high-throughput methodologies, opens new frontiers for crop improvement and the discovery of plant-derived therapeutics. Future directions will likely focus on developing even broader host-range vectors, refining tissue-specific silencing, and deepening the application of VIGS-induced epigenetics in breeding programs. For biomedical research, the principles of VIGS and the gene functions it reveals in medicinal plants provide critical insights into biosynthetic pathways, offering a foundational knowledge base for drug discovery and development. As a bridge between genomics and phenomics, VIGS will continue to be a critical technology for elucidating gene function and accelerating biotechnological innovation.
References
- 1. Virus-Induced Gene Silencing (VIGS): A Powerful Tool for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10057534/]
- 2. Applications of Virus Induced Gene Silencing (VIGS) in ... [https://www.longdom.org/open-access/applications-of-virus-induced-gene-silencing-vigs-in-plant-functional-genomics-studies-37456.html]
- 3. Virus-Induced Gene Silencing (VIGS) in Functional Genomics [https://www.mdpi.com/2311-7524/11/11/1297]
- 4. Development and application of virus-induced gene ... [https://www.sciencedirect.com/science/article/abs/pii/S030442382300763X]
- 5. A Methodological Advance of Tobacco Rattle Virus ... [https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2021.671091/full]
- 6. Truncated CMV2b N43 enhances virus-induced gene ... [https://plantmethods.biomedcentral.com/articles/10.1186/s13007-025-01446-w]
- 7. Efficient Virus-Induced Gene Silencing (VIGS) Method for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12115190/]
- 8. A long road ahead to reliable and complete medicinal plant ... [https://www.nature.com/articles/s41467-025-57448-8]
- 9. Molecular Bases of Viral RNA Targeting by Viral Small ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC1866121/]
- 10. RNA-induced silencing complex [https://en.wikipedia.org/wiki/RNA-induced_silencing_complex]
- 11. Virus-Induced Gene Silencing (VIGS): A Powerful Tool for ... [https://pubmed.ncbi.nlm.nih.gov/36982682/]
- 12. SiRNAs as antiviral drugs – Current status, therapeutic ... [https://www.sciencedirect.com/science/article/pii/S016635422400233X]
- 13. Antiviral RNA interference in plants: Increasing complexity ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12546453/]
- 14. Decay of mRNAs targeted by RISC requires XRN1, the Ski ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC1370735/]
- 15. Assessment of RNA interference effectiveness mediated by ... [https://www.sciencedirect.com/science/article/abs/pii/S0965174825001626]
- 16. Differential RNAi efficacy of siRNA and dsRNA targeting ... [https://www.frontiersin.org/journals/insect-science/articles/10.3389/finsc.2025.1574585/full]
- 17. DICER-LIKE2 Plays a Crucial Role in Rice Stripe Virus ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10675384/]
- 18. The RNA-induced Silencing Complex: A Versatile Gene ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC2709356/]
- 19. Molecular mechanisms of Dicer: endonuclease and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5415849/]
- 20. RNA-dependent RNA polymerase [https://en.wikipedia.org/wiki/RNA-dependent_RNA_polymerase]
- 21. Dicer [https://en.wikipedia.org/wiki/Dicer]
- 22. RNA Dependent RNA Polymerases: Insights from Structure ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC5850383/]
- 23. Seeking the truth behind the myth: Argonaute tales from “ ... [https://www.sciencedirect.com/science/article/pii/S1097276521009552]
- 24. Phosphorylation of human Argonaute proteins affects small ... [https://pubmed.ncbi.nlm.nih.gov/21071408/]
- 25. RNA-directed DNA methylation in plants: Where to start? [https://pmc.ncbi.nlm.nih.gov/articles/PMC3866238/]
- 26. An atypical component of RNA-directed DNA methylation ... [https://www.nature.com/articles/cr2011173]
- 27. Development of a robust and efficient virus-induced gene ... [https://plantmethods.biomedcentral.com/articles/10.1186/s13007-024-01320-1]
- 28. Selection of stable reference genes for accurate reverse- ... [https://www.nature.com/articles/s41598-025-08940-0]
- 29. Virus-Induced Gene Silencing (VIGS): A Powerful Tool for ... [https://www.mdpi.com/1422-0067/24/6/5608]
- 30. Virus-induced gene silencing: A versatile tool for discovery ... [https://www.sciencedirect.com/science/article/abs/pii/S0981942809001934]
- 31. Engineering VIGS Vectors by Modifying Movement ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11664229/]
- 32. Development of a gene silencing DNA vector derived from a ... [https://plantmethods.biomedcentral.com/articles/10.1186/1746-4811-5-9]
- 33. Cucumber mosaic virus-induced gene silencing in banana [https://www.nature.com/articles/s41598-019-47962-3]
- 34. Virus-induced gene silencing is a versatile tool for ... [https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2014.00323/full]
- 35. Applications of Virus-Induced Gene Silencing in Cotton [https://www.mdpi.com/2223-7747/13/2/272]
- 36. Recent Advances on Virus Induced Gene Silencing (VIGS) [https://www.longdom.org/open-access/recent-advances-on-virus-induced-gene-silencing-vigs-plant-functionalgenomics-33473.html]
- 37. Agroinfiltration technique for elucidating the functions of ... [https://www.nature.com/articles/s41598-025-08344-0]
- 38. Quantitative dissection of Agrobacterium T-DNA ... [https://www.nature.com/articles/s41477-025-01996-w]
- 39. Harnessing Transient Expression Systems with Plant Viral ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12193647/]
- 40. A high-throughput virus-induced gene silencing protocol ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3879149/]
- 41. Virus-induced gene silencing and its application in ... [https://www.sciencedirect.com/science/article/abs/pii/S0176161708001223]
- 42. Virus‐induced gene silencing database for phenomics and ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6508541/]
- 43. VIGS-Mediated Forward Genetics Screening for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3856292/]
- 44. Simultaneous silencing of two different Arabidopsis genes ... [https://plantmethods.biomedcentral.com/articles/10.1186/s13007-020-00701-6]
- 45. Hairpin inserts in viral genomes are stable when they ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11998532/]
- 46. A rapid virus-induced gene silencing (VIGS) method for ... [https://virologyj.biomedcentral.com/articles/10.1186/s12985-017-0716-6]
- 47. Application and Expansion of Virus-Induced Gene ... [https://www.mdpi.com/2311-7524/9/8/934]
- 48. A Cotyledon-based Virus-Induced Gene Silencing (Cotyledon ... [https://plantmethods.biomedcentral.com/articles/10.1186/s13007-024-01154-x]
- 49. A virus-induced gene silencing approach to understanding ... [https://pubmed.ncbi.nlm.nih.gov/21802100/]
- 50. Identification of DELLA and GID1 genes in Catharanthus ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12141139/]
- 51. The role of the Golden2-like (GLK) transcription factor in ... [https://link.springer.com/article/10.1007/s00299-024-03208-9]
- 52. Spray-induced gene silencing for crop protection [https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2025.1527944/full]
- 53. Comprehensive Mechanism of Gene Silencing and Its Role ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC8475493/]
- 54. Developing and applying a virus-induced gene silencing ... [https://www.sciencedirect.com/science/article/abs/pii/S0378111924009685]
- 55. California TRV-based VIGS vectors mediate gene silencing at ... [https://bmcplantbiol.biomedcentral.com/articles/10.1186/s12870-021-03324-8]
- 56. Gene Silencing and its Applications in Plants [https://www.opensciencepublications.com/fulltextarticles/JPSR-2349-2805-9-233.html]
- 57. Viral delivery of an RNA-guided genome editor for ... [https://www.nature.com/articles/s41477-025-01989-9]
- 58. New Technique Silences Plant Genes to Boost Crop Yields [https://www.seedworld.com/europe/2025/08/18/new-technique-silences-plant-genes-to-boost-crop-yields/]
- 59. Vesicular and non-vesicular extracellular small RNAs ... [https://www.nature.com/articles/s41467-025-57908-1]
- 60. An Optimized Protocol to Increase Virus-Induced Gene ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC3893464/]
- 61. Improving agroinfiltration-based transient gene expression ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6109318/]
- 62. Acetosyringone - an overview | ScienceDirect Topics [https://www.sciencedirect.com/topics/chemistry/acetosyringone]
- 63. Enhancing virus-induced gene silencing efficiency in tea ... [https://www.sciencedirect.com/science/article/abs/pii/S0304423824007398]
- 64. Refinement and Enhancement of Agrobacterium-Mediated ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC11507023/]
- 65. Advancing virus-induced gene silencing in sunflower [https://plantmethods.biomedcentral.com/articles/10.1186/s13007-024-01241-z]
- 66. VIGS-Mediated Forward Genetics Screening for Identification of Genes Involved in Nonhost Resistance - PMC [https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3856292/]
- 67. Virus-Induced Genome Editing (VIGE): One Step Away from an Agricultural Revolution [https://www.mdpi.com/1422-0067/26/10/4599]
- 68. Efficient Virus-Induced Gene Silencing in Arabidopsis - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC1557620/]
- 69. Application of RNA silencing to plant disease resistance - PMC [https://pmc.ncbi.nlm.nih.gov/articles/PMC3503840/]
- 70. Viral suppressors of RNA silencing [https://www.sciencedirect.com/science/article/abs/pii/S1360138511000458]
- 71. Truncated CMV2bN43 enhances virus-induced gene ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12542452/]
- 72. Engineering PVX vectors harboring heterologous VSRs to ... [https://pubmed.ncbi.nlm.nih.gov/41254168/]
- 73. CRISPR/Cas9 screens identify key host factors that ... [https://www.nature.com/articles/s41541-024-01007-7]
- 74. Silencing of the phytoene desaturase (PDS) gene affects ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC6777038/]
- 75. Knockout of phytoene desaturase gene using CRISPR ... [https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2022.1074541/full]
- 76. Phytoene Desaturase (PDS) Gene-Derived Markers ... [https://www.mdpi.com/2311-7524/10/3/294]
- 77. Selection of Reliable Reference Genes for ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC10379039/]
- 78. Analyzing qPCR data: Better practices to facilitate rigor and ... [https://www.sciencedirect.com/science/article/pii/S2405580825004431]
- 79. A virus-induced gene silencing (VIGS) system for functional ... [https://plantmethods.biomedcentral.com/articles/10.1186/1746-4811-10-16]
- 80. Phylogenomics and metabolic engineering reveal a ... [https://www.nature.com/articles/s41467-025-61686-1]
- 81. Recent advances in CRISPR-based functional genomics ... [https://www.nature.com/articles/s12276-024-01212-3]
- 82. Recent applications, future perspectives, and limitations of ... [https://pubmed.ncbi.nlm.nih.gov/40777740/]
- 83. State of CRISPR Gene Editing in the Drug Discovery Sector [https://www.synthego.com/blog/survey-report-crispr-therapeutics/]
- 84. CRISPR Clinical Trials: A 2025 Update [https://innovativegenomics.org/news/crispr-clinical-trials-2025/]
- 85. Virus-Induced Genome Editing (VIGE): One Step Away ... [https://pmc.ncbi.nlm.nih.gov/articles/PMC12111327/]
- 86. Systems for Targeted Silencing of Gene Expression and ... [https://www.mdpi.com/1422-0067/25/10/5231]
- 87. Efficient Virus-Induced Gene Silencing (VIGS) Method for ... [https://pubmed.ncbi.nlm.nih.gov/40431112/]
- 88. based virus-induced gene Silencing in Atriplex canescens [https://plantmethods.biomedcentral.com/articles/10.1186/s13007-025-01427-z]
- 89. Methods for effective real-time RT-PCR analysis of virus- ... [https://www.sciencedirect.com/science/article/abs/pii/S0166093406002722]
- 90. Cyclo (Pro-Tyr) upregulates GmPOD53L to enhance ... [https://www.frontiersin.org/journals/plant-science/articles/10.3389/fpls.2025.1628555/full]